
LADA Granta FL NFR R1
Шины сухие: Yokohama Advan A005 190/560-R15
Шины дождевые: Yokohama Aavan A006 190/560-R15
Рабочий объем двигателя
1600 куб.см
Мощность
155 л.с
Шасси
Ohlins
Масса
1050 кг
Тормозная система
TRW
Длина
3926
Ширина
1700
Пилоты
Ранее участвовали
Стартовый номер:
4
Класс:
Туринг-Лайт
Команда:
Стартовый номер:
8
Класс:
Туринг-Лайт
Команда:
Стартовый номер:
11
Класс:
Туринг-Лайт
Описание
ДВИГАТЕЛЬ— Максимальная мощность — 155 л. с. при 7800 об/мин
с. при 7800 об/мин
— Максимальный крутящий момент — 165 Нм при 5800 об/мин
— Дроссельная заслонка — 52 мм
— Оригинальный впускной коллектор
— Сварной выпускной коллектор
— Прямоточная система выпуска.
КПП
— 5-ти ступенчатая, синхронизированная
— Спортивные передаточные отношения, дифференциал повышенного трения дискового типа
ПОДВЕСКА
— Подрамник передней подвески
— Сварные рычаги передней подвески
— Усиленная балка задней подвески с увеличенной жесткостью
— Спортивные газонаполненные амортизаторы
ТОРМОЗНАЯ СИСТЕМА
— Задний дисковый тормоз на компонентах TRW
— Двухконтурная
— Регулятор давления в заднем контуре
КУЗОВ
— Облегченный в соответствии с техническими требованиями
Обзор Лада Калина NFR: цена, технические характеристики
Технические характеристики Лада Калина NFR
| Модельный год | 2016 |
| Тип кузова | Хэтчбек |
| Длина, мм | 3965 |
| Ширина, мм | 1700 |
| Высота, мм | 1465 |
| Количество дверей | 5 |
| Количество мест | 5 |
| Объем багажника, л | 240-550 |
| Страна сборки | Россия |
Модификации Лада Калина NFR
Лада Калина NFR 1.
 6 MT
6 MT| Максимальная скорость, км/ч | 205 |
| Время разгона до 100 км/ч, сек | 9.2 |
| Двигатель | Бензиновый |
| Рабочий объем, см3 | 1596 |
| Мощность, л.с. / оборотах | 136/6800 |
| Момент, Н·м / оборотах | 154/4750 |
| Расход комби, л на 100 км | 8.9 |
| Тип коробки передач | Механическая, 5 передач |
| Привод | Передний |
| Показать все характеристики | |
Одноклассники Лада Калина NFR по цене
К сожалению, у этой модели нет одноклассников…
Отзывы владельцев Лада Калина NFR
Лада Калина NFR, 2013 г
Шел в автосалон за Калиной спорт, а взял Лада Калина NFR. Машинка безусловно очень понравилась. И дело не только в более агрессивной внешности, она заметно динамичнее той же Калины в спортивной версии. Да, это не Ниссан ГТР или BMW M3, это ВАЗовская Калина, но мне она нравится. Подвеска у нее получилась очень жесткой, каждая кочка «аукается» в позвоночник, но зато ощущения от руления непередаваемые, какие-то ребяческие чувства просыпаются. В отличие от Калины спорт здесь чувствуется более серьезный подход к тюнингу, да и «спорта» здесь побольше. Удобные сиденья-ковши, четкая и слаженная работа коробки, отзывчивый двигатель — все это конечно очень здорово. Единственное замечание только к рулевому управлению — оно мне показалось недостаточно информативным. Машина прошла обкатку, на одометре 12 тыс. км, залил хорошее масло, в планах поставить турбину, ну это пока только в планах.
Машинка безусловно очень понравилась. И дело не только в более агрессивной внешности, она заметно динамичнее той же Калины в спортивной версии. Да, это не Ниссан ГТР или BMW M3, это ВАЗовская Калина, но мне она нравится. Подвеска у нее получилась очень жесткой, каждая кочка «аукается» в позвоночник, но зато ощущения от руления непередаваемые, какие-то ребяческие чувства просыпаются. В отличие от Калины спорт здесь чувствуется более серьезный подход к тюнингу, да и «спорта» здесь побольше. Удобные сиденья-ковши, четкая и слаженная работа коробки, отзывчивый двигатель — все это конечно очень здорово. Единственное замечание только к рулевому управлению — оно мне показалось недостаточно информативным. Машина прошла обкатку, на одометре 12 тыс. км, залил хорошее масло, в планах поставить турбину, ну это пока только в планах.
Достоинства: отзывчивый двигатель. Собранная подвеска.
Недостатки: неинформативный руль.
Владислав, Санкт-Петербург
Обои рабочего стола Лада Калина NFR
1024×768
1280×1024
Обои Лада Калина NFR
Lada Kalina NFR / Лада Калина NFR
Калина NFR (Need for Race, что обозначается как «жажда гонки») — самая «спортивная» Лада в семействе ВАЗ. Глядя на рельефную внешность Лада Калина NFR нельзя не отметить мускулистые обтекатели порогов и бампера, которые формируют незабываемый образ уверенного в себе атлета. 17-дюймовые литые колеса с 7-спицевым дизайном, а также аккуратно заниженный профиль выдают в этом автомобиле неистощимую энергичность и твердый характер. Компактное заднее антикрыло, как завершающий штрих, эффектно дополняет целостность образа. Сиденья в Lada Kalina NFR выполнены из мягких нескользящих материалов и имеют ярко-выраженную поддержку тела. Расставленные по салону мелкие акценты в виде надписей NFR, контрастной прострочки, а также руль со спортивным удобным хватом намекают на эмоциональный характер автомобиля и его гоночные способности.
Глядя на рельефную внешность Лада Калина NFR нельзя не отметить мускулистые обтекатели порогов и бампера, которые формируют незабываемый образ уверенного в себе атлета. 17-дюймовые литые колеса с 7-спицевым дизайном, а также аккуратно заниженный профиль выдают в этом автомобиле неистощимую энергичность и твердый характер. Компактное заднее антикрыло, как завершающий штрих, эффектно дополняет целостность образа. Сиденья в Lada Kalina NFR выполнены из мягких нескользящих материалов и имеют ярко-выраженную поддержку тела. Расставленные по салону мелкие акценты в виде надписей NFR, контрастной прострочки, а также руль со спортивным удобным хватом намекают на эмоциональный характер автомобиля и его гоночные способности.
Установленный на подрамник, двигатель на Лада Калина NFR подвергся небольшой доработке систем впуска и выпуска, теперь он выдает 136 сил, что позволяет развивать максималку на уровне 203 км/час. Изменения произошли также и в подвеске хэтчбека, но компоновка шасси осталась прежней. Спереди стоят все те же стойки МакФерсон, сзади — полузависимая балка. Чтобы побороть крены последнюю сделали еще более жесткой, плюс по всему периметру установили иные амортизаторы. Такой комплекс мер позволил основательно улучшить управляемость и маневренность автомобиля, особенно в условиях агрессивной езды. Единственная комплектация Лада Калина NFR представлена аудиосистемой с сенсорным управлением, Bluetooth с опцией «свободные руки», климатическим контролем, датчиком дождя, электрообогревом зеркал. Также на автомобиль установлены дневные ходовые огни.
Спереди стоят все те же стойки МакФерсон, сзади — полузависимая балка. Чтобы побороть крены последнюю сделали еще более жесткой, плюс по всему периметру установили иные амортизаторы. Такой комплекс мер позволил основательно улучшить управляемость и маневренность автомобиля, особенно в условиях агрессивной езды. Единственная комплектация Лада Калина NFR представлена аудиосистемой с сенсорным управлением, Bluetooth с опцией «свободные руки», климатическим контролем, датчиком дождя, электрообогревом зеркал. Также на автомобиль установлены дневные ходовые огни.
Калина for Race | Русский Автомобиль
Need for Race – так расшифровывается аббревиатура NFR. Предположение Авторевю о том, что мощная Калина будет обозначаться как GT не оправдалось. Этой машины официально пока нет, никто не назовёт её характеристик и уж тем более цен и комплектаций. Это незавершённый проект, постоянно дорабатываемый.
Серийные образцы появятся, дай бог, только к концу года. Но у нас есть возможность познакомиться с Lada Kalina NFR уже сейчас.
Внешне NFR почти полностью повторяет Калину Спорт, презентация которой состоится 7 июня. Вот для сравнения обе модели рядом:
Обвес, сделанный Андреем Рузановым, одинаков, за исключением двух вентиляционных отверстий на крышке капота у NFR. Это такая «фишка», по которой можно узнать самую мощную Калину. Но… Почему-то мне кажется, что в процессе подготовки производства от этих дырок откажутся – нетехнологично, почти бессмысленно с точки зрения охлаждения (а с точки зрения запыливания моторного отсека – так и просто вредно). Хотя выглядит эффектно.
В любом случае, NFR можно будет узнать по другим колёсам – семнадцатидюймовым (у Калины Спорт – 16 дюймов), с пятью крепёжными болтами вместо четырёх.
В салоне отличий поболее. Главное – для Калины NFR сделали другие передние сиденья, с более развитой боковой поддержкой. Они обшиты кожей, а прострочка сделана жёлто-оранжевой (вместо красного у ГС/КС).
Появилась даже оригинальная эмблема модели, вышитая на спинках кресел.
Этот красивый жёлтый шов везде. На всех тканевых и кожаных элементах.
Руль тоже «кожаный», с нарочито гоночной риской нулевого положения.
В остальном – Калина как Калина. Если, конечно, не знать, «что у ей в нутре».
А там – форсированный до 135 л.с двигатель 21126. Главное изменение – впускной ресивер, конструктивно похожий на то, что используется в кубковых Грантах. Новый впуск позволил поднять крутящий момент на средних оборотах, в районе 2000 – 3000 об. и выровнять тяговую характеристику во всём диапазоне. Двигатель сейчас реально «крутится» до 7000 об. Распредвалы тоже оригинальные, ещё более «жёсткие», чем на 120-сильном двигателе.
Выпуск тоже переработали. Причём существенно – изменили сам выпускной коллектор, улучив продувку цилиндров. Нейтрализатор остался штатным, а после него – система глушителей от текущей версии Гранты Спорт. Ну, почти.
Калибровка двигателя заняла несколько месяцев и работа ещё отнюдь не закончена, ещё будут вносить изменения в программное обеспечение.
Двигатель вкупе с передней подвеской установлены на подрамник собственной конструкции. Он крепится к штатным точкам «калиновского» кузова. Подрамник этот пригоден не только для дорожной версии машины, но и для спорта, в машинах, подготовленных по группе R2b (как «лукойловские» Калины из Туринг-лайт, например). Приводы колёс оригинальные – так как используются ступицы от Renault, то и валы пришлось взять соответствующие. Внешние ШРУСы – французские, внутренние – ВАЗовские.
Коробка передач останется от Калины, с главной парой 4,3. Её входной момент находится на пределе по отношению к 135-сильному двигателю. Или даже немножко за – абсолютного показателя крутящего момента создатели пока не приводят. Как не приводят и динамико-скоростных характеристик; всё, дескать, пока в доработке. А вот с самоблокирующимся дифференциалом определились однозначно – его не будет.
Тормозные диски увеличились в диаметре, до 282 мм. Вообще, почти все основные элементы тормозной системы взяты от Renault Megane с двухлитровым двигателем (суппорты, главный тормозной цилиндр и пр. ).
).
Задняя подвеска по архитектуре осталась «калиновской», только усилили поперечную балку. Уменьшили толщину проставки. Пружины и амортизаторы оригинальные, сделанные на основе тех, что сейчас используются для Гранты Спорт. Не исключено, что их унифицируют (то есть тупо возьмут грантовские). Пока же Kalina NFR «сидит пониже» и имеет меньший ход подвески.
Конечно, системы стабилизации в этом автомобиле не будет – адаптация заняла бы слишком много времени (и денег, что немаловажно). А с ABS научились справляться ещё на предыдущей модели, делая специальные проточки в задних ступицах, витруально конвертирующих барабанные тормоза в дисковые. На эффективность торможения это не влияет, сам проверял неоднократно на дромовской ГС.
«Прострелы» по щербатому тольяттинскому асфальту на Kalina NFR – тот ещё экспириенс. Тяга ровная до предельных 7000 об, и даже на пределе можно ехать довольно эффективно, благо, электронного ограничителя тут нет – не успели поставить. Нет никакой болтанки и дрожания момента, всё предельно стабильно. Остаётся только уповать на собственное здравомыслие и не перекрутить мотор.
Остаётся только уповать на собственное здравомыслие и не перекрутить мотор.
На низких оборотах ведёт себя примерно так же, как Гранта Спорт: в самом низу есть провальчик, а потом сразу обозначается подхват. Провал компенсируется растянутой характеристикой включения сцепления: педаль длинноходная, тронутся можно очень плавно. Нет резкости при старте, характерной для спортивных сцеплений, с их жёсткими прижимающими пружинами.
Конечно, шумно. И двигатель голосит, и спрямлённый выпускной тракт… Если над акустикой не поработают, то в правила ЕЭК ООН по внутреннему шуму машина точно не впишется. Хотя мне характер звучания этой машины нравится, он более гоночный, чем на Гранте Спорт.
Глобальная проблема – рулевое. При такой настройке шасси оно уже выглядит чужеродно. Если на базовой Калине рейка с таким передаточным отношением вполне гармонична, тот тут – вызывает недоумение. Хочется не столько даже точности руления, с ним всё более-менее в порядке, подрамник сыграл свою роль. Хочется более быстрого отклика на руль, без необходимости на него наматываться. На змейке я понял, что не хочу ехать по змейке, руль приходится вращать так много, что суставы начинают просить прощения.
Хочется более быстрого отклика на руль, без необходимости на него наматываться. На змейке я понял, что не хочу ехать по змейке, руль приходится вращать так много, что суставы начинают просить прощения.
Колёса задевают за арки в крайних положениях и с этим что-то надо делать. Но что тут сделаешь, если 17 дюймов для этой модели – принципиальный момент? Виталий Прилак, замдиректора Лады Спорт обещал «учесть и разобраться».
Подвеска жёсткая, даже очень. Каждая пылинка на дороге отдаётся встряхиванием. Ехать по Тольятти – мука, постоянно боишься пробить колесо. Профиль шин настолько мал, что демпфирующих функций резина почти не выполняет. Такие шины в наших условиях будут расходным материалом. В Москве ещё можно ездить, но в провинции, с её дорожной неухоженностью я бы два раза подумал, прежде чем покупать такой болид.
Очень понравились передние кресла, боковая поддержка там реализована ровно в той области спины, в какой нужно. Во всяком случае, для человека моего роста. И садиться удобно, и в поворотах не сползаешь наружу.
Позволю себе небольшое обобщение.
Чем больше дорабатываешь серийный автомобиль, тем больше начинают мешать особенности, которые доработать не можешь.
В данном случае это изначально высокая «калиновская» посадка и верхнее крепление рулевого механизма (на подрамник его не перенесёшь, хоть убейся). Отличное шасси предельно диссонирует с допотопными решениями по посадке и рулевому. В Гранте Спорт это не так бросается в глаза, уровень претензий там поменьше. А в NFR уже есть замах на участие в реальных гонках. И в этом месте концепция автомобиля рушится, под натиском системных противоречий.
Kalina NFR явно получилась машиной не для всех. Возможную розничную цену мне не смогли назвать даже в первом приближении, сказали только, что «в 600 тыс не впишемся». Ну, пусть будет 650 тыс руб, за хорошую комплектацию. Конкурентов всё равно в этом ценовом диапазоне нет. Ближайший по идеологии автомобиль – новый Renault Clio RS – стоит больше миллиона. Конечно, есть хэтчбеки В- и С-классов с моторами аналогичной мощности, но они не спортивные по самому духу, там и подвеска «гражданская», и кузова много тяжелее. А NFR – это чистая железка, без электроники, дающая скромной энерговооружённостью столько ощущений, сколько не дадут ни Opel Astra OPC, ни Volkswagen Golf GTI.
А NFR – это чистая железка, без электроники, дающая скромной энерговооружённостью столько ощущений, сколько не дадут ни Opel Astra OPC, ни Volkswagen Golf GTI.
Даёшь новый монокубок?
Нравится(2)Не нравится(0)
При использовании материала, пожалуйста, сделайте ссылку на Русский автомобиль.
| OEM номер | 21925-2215010-37 |
| OEM номер | 21925-2215010-37 21925-2215008-81 |
| OEM номер | 21925-2215010-37 21925-2215008-81 |
| Ось (сторона установки) | передняя правая |
| Твердость материала | HB 10-1=248Мпа |
| Марка стали | 55 марка, Углеродистая |
| Диаметр вала,мм |
27. 5 5
|
| Длинна, мм. | 884 |
| Количество зубьев | 25 |
| Количество зубьев наружного шарнира | 25 |
| Количество зубьев внутреннего шарнира | 24 |
«Я буду настойчив»: Шейн Проктор, лучший наездник Вашингтона, борется за изнурительный физический и эмоциональный год за 6-й NFR
Кивок головы, и ворота парашюта захлопываются.
Сила быка весом почти в 1 тонну врывается на арену, копыта бьют по земле, пока Шейн Проктор держится одной рукой, а другую держит в воздухе.
После возможно самых длинных 8 секунд в спорте раздается звуковой сигнал.
После этих 8 секунд у ковбоя родео Гранд-Кули, штат Вашингтон, остается мало времени, чтобы беспокоиться о том, что пошло правильно, а что нет.
На трассе родео ждет еще один бык в других воротах в другом штате.
В этом году Проктору повезло меньше. Было больное плечо после того, как его оторвали на одном родео. И надоедливая травма паха, и недостаток сна.
Хуже всего было, когда бык наступил ему на лодыжку.
Проктор, цитируя свой любимый фильм «8 секунд», сказал: «Говорят, дело не в том, когда тебя ранят в езде на быке, а в том, насколько это плохо».
Всегда трудно попасть на национальный финал Родео, но этот год был особенно трудным для 36-летнего Проктора, чемпиона Ассоциации профессиональных ковбоев родео (PRCA) 2011 года по верховой езде на быках, который родился и вырос в Гранд-Кули и является чемпионом штата. лучший спортсмен родео.
Сезон 2021 года для Проктора начался в ноябре 2020 года, через месяц после завершения регулярного сезона 2020 года, в Остине, штат Техас, где он набрал 90 очков и занял второе место. Несмотря на то, что хорошая езда и место в соревнованиях стали отличным началом сезона, за это пришлось заплатить.
«В конце концов, я снова испортил TCL в правом колене, который я испортил четыре года назад и отказался от операции», — сказал Проктор. «Но это произошло в действительно хорошее время, потому что это не было сердцем сезона и было еще до того, как все началось.
Необычное время простоя также дало Проктору возможность побыть со своей женой Хейли и дочерью Кули Мэй Проктор, которая родилась всего пять дней спустя.
После насыщенного событиями старта Проктор принял участие в зимнем родео в помещении в Техасе и в «Калифорнийском забеге» весной, где ему предстояло одержать крупную победу в Ред-Блаффе, событии, которое он всегда хотел выиграть за свои 17 лет профессиональной езды на быках. .
.
По мере приближения напряженного (и потенциально чрезвычайно прибыльного) лета сезона Проктор разогревался и оказался на 13-м месте в турнирной таблице и по восхождениям.Но июньской ночью в Понка-Сити, штат Оклахома, все изменилось.
Проктор оседлал своего быка, набрав 85,50 балла и заняв четвертое место, но поездка также привела к перелому ноги. Он сломал правую малоберцовую кость в трех местах, и хирургам пришлось вставить пластину и три винта.
«После этого мне пришлось пропустить забег 4 июля, и врачи сказали мне, что пройдет три месяца, прежде чем я смогу кататься, потому что у меня были проблемы с сухожилиями в лодыжке, когда я сломал ногу», — сказал Проктор.
Три месяца перерыва в самом сердце сезона угрожали положить конец любому его шансу на успех в финале родео, где квалифицируются 15 лучших заработков по итогам года. Сезон Проктора мог закончиться прямо сейчас.
Он отказался уходить. Родео принес ему более миллиона долларов. Он пять раз выходил в финал и выигрывал отдельный чемпионат Ассоциации профессиональных родео-ковбоев.
«Мое мнение о моей ноге и о том, что я поправилась на два месяца раньше, заключалось в том, что это не так больно, как я думал, — сказал Проктор.«Было больно, но это было не то, что было у меня в голове. И это была моя философия. Мой старый тренер по родео (в Северо-Западном колледже в Пауэлле, штат Вайоминг) Гэвин Глайх однажды спросил меня: «Ты ранен или тебе больно?» Если вы ранены, вы можете ездить, если вы ранены, вы не можете». В этом году я немного раздвинул границы», — сказал он со смешком.
Получив приглашение на престижный турнир Calgary Stampede, Проктор снял гипс через месяц после операции и направился на север, в Канаду.
К тому времени, когда в конце июля начались Дни границы Шайенна в столице Вайоминга, Проктор опустился на 17 позиций до 30-го, и пришло время немного изменить его.
«В тот момент это было просто для того, чтобы доказать всем и себе, что я могу сделать это, несмотря ни на что», — сказал Проктор. «Я многое изменил в этом сезоне, основываясь на этом заключении; Я вошел немного по-другому, мне пришлось перейти на ботинки для верховой езды, которые должны были шнуроваться, мне пришлось носить подушечки на лодыжке, временами это казалось больше дракой, чем веселой поездкой, но я хотел доказать, что я могу это сделать себе больше всего на свете.
Проктор отправился в путь, как и любой другой август, зарабатывая солидные поездки и деньги по стране, а также победу в Линдене, штат Вашингтон.
В сентябре, когда до конца сезона оставался месяц, Северо-Западный пробег привел Проктора домой.
На соревнованиях Ellensburg Xtreme Bulls он набрал 88 очков и занял второе место в раунде, а также занял третье и четвертое место в общем зачете, но Проктор порвал обе вращательные манжеты, и его хирургически восстановленное левое плечо сместилось.
«Это расстраивает, когда этим летом я ехал только с болью. От сломанной ноги до этого. Но я буду настойчив», — написал Проктор в своем посте в Instagram.
«В этом году я узнал о себе ментально и физически больше, чем в любой другой год, — сказал Проктор. «Это определенно было самым трудным и упорным из того, что мне приходилось делать. У меня и раньше были проблемы с плечами, но за этот год я действительно многое узнал о себе и о том, с чем я могу мириться».
На финале в Лас-Вегасе Проктор оседлал своих первых двух быков.Затем вспыхнули травмы.
«Это морально утомительно, потому что иногда начинаешь сомневаться в себе, а на самом деле этот вид спорта во многом связан с уверенностью», — сказал Проктор. «Куда бы мы ни пошли, мы платим за наш собственный бензин, нашу собственную еду, наши вступительные взносы и все, что нам нужно, чтобы двигаться дальше, поэтому вы, по сути, каждый раз играете на себя, чтобы делать добро и побеждать. И тот факт, что вы устали от путешествий и этих 18-часовых поездок всю ночь, затем езда на быках, выходы из дома и снова, эти долгие ночи требуют много размышлений про себя и выяснения того, из чего вы сделаны.
Бесчисленные проблемы могут разочаровывать и обескураживать, но у Проктора есть большой круг сторонников в его углу, его семья, которая всегда старалась поддержать его дух и помочь ему в дороге, например, его отец Лаки, который водит машину. его сын по всей трассе родео, когда Проктору нужен перерыв за рулем.
«Мои родители всегда беспокоились обо мне, когда я в дороге, но они также всегда были моими поклонниками № 1 и самыми большими сторонниками, моя жена — то же самое», — сказал Проктор.«Они всегда мотивируют меня и подталкивают меня стать лучше».
И хотя национальный финал Родео в этом году, возможно, прошел не так, как надеялся Проктор, его дочь Кули была самым значимым аспектом мероприятия.
«Видеть мою дочь на сцене, когда мы получили наши последние номера, и просто видеть ее там, чтобы увидеть и испытать то, что делает ее отец, хотя я знаю, что она этого не запомнит, но у нас будут фотографии, чтобы показать ей, насколько она была особенной. для нас там, — сказал Проктор.
Семьятакже является причиной того, что Проктор надеется немного отойти от верховой езды в 2022 году и в основном следовать за своей женой Хейли, которая выступает со своими лошадьми на родео по всей стране и соревнуется там, где она выступает.
«Катание на быках — очень эгоистичный вид спорта; мы проводим так много времени, просто идя вверх и вниз по дороге, думая о себе, и после того, как у меня родилась маленькая девочка, я больше этого не делал», — сказал Проктор. «Наша маленькая девочка побывала в 22 штатах уже в первый год своей жизни, и это было потрясающе — возить ее в эти места, но в то же время было душераздирающе быть вдали от нее.В середине лета я решил, что не хочу ложиться спать с Кули и не быть рядом, когда она проснется, что мне приходилось делать пару раз в этом году».
«Наша маленькая девочка побывала в 22 штатах уже в первый год своей жизни, и это было потрясающе — возить ее в эти места, но в то же время было душераздирающе быть вдали от нее.В середине лета я решил, что не хочу ложиться спать с Кули и не быть рядом, когда она проснется, что мне приходилось делать пару раз в этом году».
Тем не менее, Проктор благодарен, что смог вырастить Кули вокруг родео, спорта, который дал ему «все в жизни», от оплаты обучения в колледже, титула чемпиона, путешествий по стране до встречи с женой и девочка.
Жизнь и год, определяемые более чем 8 секундами за один раз, взятые за один день, одну милю и одного быка за один раз.
— Мне нечего доказывать, — сказал Проктор.
Генетически кодируемый одноволновой датчик для визуализации цитозольной и клеточной поверхности АТФ
Реферат
Аденозин-5′-трифосфат (АТФ) является универсальным внутриклеточным источником энергии и эволюционно древним, вездесущим внеклеточным сигналом у различных видов. Здесь мы сообщаем о создании и характеристике одноволновых генетически кодируемых флуоресцентных сенсоров (iATPSnFR) для визуализации внеклеточной и цитозольной АТФ путем вставки циклически переставленной суперпапки GFP в эпсилон-субъединицу F 0 F 1 -АТФазы из Bacillus PS3 .На поверхности клетки и в цитозоле iATPSnFR 1,0 реагирует на соответствующие концентрации АТФ (от 30 мкМ до 3 мМ) быстрым увеличением флуоресценции. iATPSnFR могут быть генетически нацелены на определенные типы клеток и субклеточные компартменты, отображаемые с помощью стандартных световых микроскопов, не реагируют на другие нуклеотиды и нуклеозиды и при слиянии с красным флуоресцентным белком функционируют как логометрические индикаторы. После тщательного рассмотрения их умеренной чувствительности к pH iATPSnFRs представляют собой многообещающие реагенты для визуализации АТФ во внеклеточном пространстве и внутри клеток в различных условиях, а также для дальнейших уточнений для конкретных приложений.
Здесь мы сообщаем о создании и характеристике одноволновых генетически кодируемых флуоресцентных сенсоров (iATPSnFR) для визуализации внеклеточной и цитозольной АТФ путем вставки циклически переставленной суперпапки GFP в эпсилон-субъединицу F 0 F 1 -АТФазы из Bacillus PS3 .На поверхности клетки и в цитозоле iATPSnFR 1,0 реагирует на соответствующие концентрации АТФ (от 30 мкМ до 3 мМ) быстрым увеличением флуоресценции. iATPSnFR могут быть генетически нацелены на определенные типы клеток и субклеточные компартменты, отображаемые с помощью стандартных световых микроскопов, не реагируют на другие нуклеотиды и нуклеозиды и при слиянии с красным флуоресцентным белком функционируют как логометрические индикаторы. После тщательного рассмотрения их умеренной чувствительности к pH iATPSnFRs представляют собой многообещающие реагенты для визуализации АТФ во внеклеточном пространстве и внутри клеток в различных условиях, а также для дальнейших уточнений для конкретных приложений.
Введение
ВВЕДЕНИЕ
Приведенные широко распространенные роли в энергетическом гомеостазе и сигнализации ячейки 1 — 5 , несколько методов были использованы для обнаружения АТФ с использованием химических подходов малой молекулы 6 , Firefly Luciferase 7 8 8 , ионный канал-экспрессирующий «Снифферные» клетки 9 — 11 — 11 , Voltammetry 12 и различные типы микроэлектродов 13 , 14 .Несмотря на достигнутый прогресс, этим методам, однако, не хватает пространственного разрешения, их нельзя легко использовать на срезах тканей или in vivo, они реагируют на нецелевые лиганды, нуждаются в чувствительных камерах для подсчета фотонов, являются неточными и/или неизбежно повреждают ткань во время исследования. размещение зонда. Важно отметить, что некоторые из этих методов не могут быть генетически нацелены на конкретные клетки, субклеточные компартменты или целые организмы, и всем им не хватает пространственного разрешения в клеточном масштабе. Хотя люцифераза генетически кодируется 15 , она требует экзогенного субстрата люциферина, добавление которого усложняет использование в срезах тканей и in vivo.Он также насыщается при наномолярном АТФ, что ниже, чем концентрации, ожидаемые во время передачи сигналов АТФ (от ~ 1 мкМ до ~ 1 мМ), и поэтому может наиболее надежно реагировать как индикатор ступенчатой функции. Кроме того, биолюминесцентный выход люциферазы обеспечивает низкие потоки фотонов и ограничивает визуализацию с разрешением в клеточном масштабе. Несмотря на эти потенциальные ограничения, в нескольких новаторских исследованиях успешно использовались генетически направленные зонды на основе люциферазы для визуализации динамики АТФ, и это остается полезным подходом в области пуринергической передачи сигналов и внутриклеточного гомеостаза АТФ 16 – 19 .Однако для удовлетворения вышеупомянутых ограничений было разработано более широко, генетически кодированные флуоресцентные датчики ATP, включая Perceval и Percevalhr 20 , 21 , ATEAM 22 и Queen 23 .
Хотя люцифераза генетически кодируется 15 , она требует экзогенного субстрата люциферина, добавление которого усложняет использование в срезах тканей и in vivo.Он также насыщается при наномолярном АТФ, что ниже, чем концентрации, ожидаемые во время передачи сигналов АТФ (от ~ 1 мкМ до ~ 1 мМ), и поэтому может наиболее надежно реагировать как индикатор ступенчатой функции. Кроме того, биолюминесцентный выход люциферазы обеспечивает низкие потоки фотонов и ограничивает визуализацию с разрешением в клеточном масштабе. Несмотря на эти потенциальные ограничения, в нескольких новаторских исследованиях успешно использовались генетически направленные зонды на основе люциферазы для визуализации динамики АТФ, и это остается полезным подходом в области пуринергической передачи сигналов и внутриклеточного гомеостаза АТФ 16 – 19 .Однако для удовлетворения вышеупомянутых ограничений было разработано более широко, генетически кодированные флуоресцентные датчики ATP, включая Perceval и Percevalhr 20 , 21 , ATEAM 22 и Queen 23 . Эти датчики ценны, но они также имеют некоторые ограничения. В частности, они либо реагируют на соотношение АДФ/АТФ вместо концентрации АТФ, либо подвержены оптическому перекрытию с клеточными источниками аутофлуоресценции, а некоторые несовместимы с одноволновой флуоресцентной визуализацией, рабочей лошадкой функциональной флуоресцентной микроскопии 24 .
Эти датчики ценны, но они также имеют некоторые ограничения. В частности, они либо реагируют на соотношение АДФ/АТФ вместо концентрации АТФ, либо подвержены оптическому перекрытию с клеточными источниками аутофлуоресценции, а некоторые несовместимы с одноволновой флуоресцентной визуализацией, рабочей лошадкой функциональной флуоресцентной микроскопии 24 .
В настоящем исследовании мы ставили перед собой цели (1) разработать датчик АТФ, который можно было бы использовать для рутинной флуоресцентной визуализации, например, при возбуждении GFP на длине волны 488 нм и испускании ~525 нм, и (2) разместить такой датчик на клетке. поверхности и внутри клеток для изображения АТФ. QUEEN — датчик коэффициента возбуждения, а ATeam — датчик флуоресцентного резонансного переноса энергии (FRET). Эти методы требуют специального оборудования, существенно замедляют скорость визуализации, усложняют использование с другими реагентами и часто имеют более низкое отношение сигнал/шум (SNR). ), чем отдельные индикаторы интенсивности флуоресценции.Кроме того, хотя индикаторы на основе FRET можно использовать в одноволновом режиме путем возбуждения на пике возбуждения донора и измерения сенсибилизированного излучения акцептора, такие стратегии не могут извлечь выгоду из логометрической природы FRET и проблематичны для количественной оценки с помощью одноволновой визуализации. 25 . Существующие датчики ATP на основе FRET также используют FRET CFP/YFP, который потребует возбуждения на 442 нм для визуализации излучения YFP: лазеры с длиной волны 442 нм не являются стандартными для конфокальных микроскопов, а CFP в существующих датчиках имеет один из самых низких квантовых выходов среди доступных. Белки на основе GFP.Это привело бы к неэффективной передаче энергии от возбужденного состояния донора к акцептору за счет диполь-дипольного взаимодействия. Напротив, наше внимание к разработке датчика АТФ на основе GFP основано на достижениях, достигнутых с поколением датчиков глутамата на основе GFP, которые являются надежной рабочей лошадкой в различных приложениях 26 , 27 .
), чем отдельные индикаторы интенсивности флуоресценции.Кроме того, хотя индикаторы на основе FRET можно использовать в одноволновом режиме путем возбуждения на пике возбуждения донора и измерения сенсибилизированного излучения акцептора, такие стратегии не могут извлечь выгоду из логометрической природы FRET и проблематичны для количественной оценки с помощью одноволновой визуализации. 25 . Существующие датчики ATP на основе FRET также используют FRET CFP/YFP, который потребует возбуждения на 442 нм для визуализации излучения YFP: лазеры с длиной волны 442 нм не являются стандартными для конфокальных микроскопов, а CFP в существующих датчиках имеет один из самых низких квантовых выходов среди доступных. Белки на основе GFP.Это привело бы к неэффективной передаче энергии от возбужденного состояния донора к акцептору за счет диполь-дипольного взаимодействия. Напротив, наше внимание к разработке датчика АТФ на основе GFP основано на достижениях, достигнутых с поколением датчиков глутамата на основе GFP, которые являются надежной рабочей лошадкой в различных приложениях 26 , 27 . Кроме того, недавно были созданы датчики на основе GFP для различных малых молекул, таких как дофамин, ацетилхолин и ГАМК 28 – 31 .Наш подход к использованию GFP, конечно, не наносит ущерба другим вышеупомянутым подходам, таким как дальнейшее использование дикого типа или модифицированных люцифераз 16 – 19 , 32 . С этой точки зрения наш подход дополняет друг друга, и мы считаем, что его легче интегрировать в стандартные режимы визуализации для визуализации в клеточном масштабе в различных биологических приложениях в клетках и в естественных условиях. Таким образом, мы сообщаем о генетически кодируемом одноволновом датчике на основе GFP для визуализации цитозольной и клеточной поверхности АТФ.
Кроме того, недавно были созданы датчики на основе GFP для различных малых молекул, таких как дофамин, ацетилхолин и ГАМК 28 – 31 .Наш подход к использованию GFP, конечно, не наносит ущерба другим вышеупомянутым подходам, таким как дальнейшее использование дикого типа или модифицированных люцифераз 16 – 19 , 32 . С этой точки зрения наш подход дополняет друг друга, и мы считаем, что его легче интегрировать в стандартные режимы визуализации для визуализации в клеточном масштабе в различных биологических приложениях в клетках и в естественных условиях. Таким образом, мы сообщаем о генетически кодируемом одноволновом датчике на основе GFP для визуализации цитозольной и клеточной поверхности АТФ.
Результаты
Создание одноволновых датчиков АТФ
Мы рассмотрели и первоначально исследовали различные потенциальные методы создания датчика АТФ. После безуспешных попыток модифицировать АТФ-управляемые Р2Х-рецепторы 33 и бактериальный регуляторный белок GlnK1 (как в Персевале 20 ) под наши нужды, мы обратились к микробным F 0 F 4 — 90 субблоки synthase epsilon 34 , которые были адаптированы для создания датчика FRET ATeam 22 и датчика степени возбуждения QUEEN 22 , 23 , которые основаны на наших датчиках 0,
. Предполагается, что субъединица эпсилон претерпевает большие конформационные изменения 34 при связывании АТФ (рис. ), а гомолог из термофильной бактерии Bacillus PS3 демонстрирует соответствующую чувствительность к связыванию АТФ 35 . Эпсилон-субъединица состоит из восьми N-концевых β-цепей, за которыми следуют две С-концевые α-спирали (рис. ), которые отходят от β-цепей в отсутствие АТФ, но при связывании колыбельной АТФ против β-цепей .
Предполагается, что субъединица эпсилон претерпевает большие конформационные изменения 34 при связывании АТФ (рис. ), а гомолог из термофильной бактерии Bacillus PS3 демонстрирует соответствующую чувствительность к связыванию АТФ 35 . Эпсилон-субъединица состоит из восьми N-концевых β-цепей, за которыми следуют две С-концевые α-спирали (рис. ), которые отходят от β-цепей в отсутствие АТФ, но при связывании колыбельной АТФ против β-цепей . Разработка и оптимизация одноволнового датчика АТФ. a Схема, показывающая конструкцию и рабочий процесс, использованные для оптимизации QUEEN-7µ в датчик АТФ с одной длиной волны с целью отображения датчика на поверхности клеток. b Кривые доза-реакция iATPSnFR в течение нескольких последовательных раундов мутагенеза (Ex: 488 нм, Em: 515 нм). Тушение флуоресценции при очень высоких концентрациях АТФ можно наблюдать в дополнение к зависимому от связывания увеличению. c Кривые доза-реакция для очищенного ATeam, QUEEN-7µ, iATPSnFR 1.0 и iATPSnFR 1.1 . Кривые доза-реакция ATeam были получены с Ex: 435 нм и Em: 530 нм. Другие конструкции были с Ex: 488 нм, Em: 515 нм. d Кривые доза-реакция очищенного iATPSnFR 1,1 на АТФ, АДФ, АМФ и аденозин. e , f Спектры возбуждения и испускания для iATPSnFR 1,0 и iATPSnFR 1,1 в растворе (кривые представляют собой среднее из 48 повторов каждого в 96-луночном планшете). Столбики погрешностей представляют s.Эм. а в некоторых случаях меньше, чем символы, используемые для среднего значения. Когда больше одного (в случае образцов трасс и графиков), n предоставляется на панелях рисунка и относится к количеству независимых оценок
c Кривые доза-реакция для очищенного ATeam, QUEEN-7µ, iATPSnFR 1.0 и iATPSnFR 1.1 . Кривые доза-реакция ATeam были получены с Ex: 435 нм и Em: 530 нм. Другие конструкции были с Ex: 488 нм, Em: 515 нм. d Кривые доза-реакция очищенного iATPSnFR 1,1 на АТФ, АДФ, АМФ и аденозин. e , f Спектры возбуждения и испускания для iATPSnFR 1,0 и iATPSnFR 1,1 в растворе (кривые представляют собой среднее из 48 повторов каждого в 96-луночном планшете). Столбики погрешностей представляют s.Эм. а в некоторых случаях меньше, чем символы, используемые для среднего значения. Когда больше одного (в случае образцов трасс и графиков), n предоставляется на панелях рисунка и относится к количеству независимых оценок
Циклически переставлено (cpGFP) 36 было вставлено между двумя α -спирали эпсилон-субъединицы после остатка 107 с расчетом на то, что изменение конформации эпсилон-субъединицы может изменить флуоресценцию. Первый линкер (L1) изначально состоял из Thr-Arg, а второй линкер (L2) из Leu-Gly (рис.). Основываясь на нашем прошлом опыте с глутаматным сенсором iGluSnFR 26 , 27 , мы начали мутировать остатки в линкерах и провели скрининг ~8500 колоний для разработки сенсоров с большим АТФ-зависимым увеличением интенсивности флуоресценции (d F / Ф ). Лучший вариант из этого скрининга имел большое изменение флуоресценции (максимум d F / F ~3,9). Однако он не смог экспрессироваться на поверхности клеток HEK293 при клонировании в вектор экспрессии млекопитающих pDisplay, который использует сигнал секреции IgG и трансмембранный домен рецептора тромбоцитарного фактора роста (PDGFR) для закрепления его на мембране.Мы пришли к выводу, что более стабильная форма GFP может улучшить сворачивание и трафик, и поэтому клонировали суперпапку с круговой перестановкой GFP 36 (cpSFGFP) вместо cpGFP. Замена cpGFP на cpSFGFP исправила поверхностный перенос клеток HEK293 (см. следующий раздел), но значительно уменьшила вызванные АТФ изменения флуоресценции. Чтобы исправить это, мы повторно оптимизировали L1 и L2 для конструкции cpSFGFP, мутировав аминокислоты в линкерах и слегка изменив их длину; Было проверено около 7000 колоний (рис.). Мы также мутировали аминокислоты (Thr9Val и Asn78Tyr), предсказанные на основе молекулярного моделирования, для уменьшения образования димеров. Благодаря этому процессу мы разработали два датчика, которые демонстрировали значительное АТФ-зависимое увеличение флуоресценции (рис. ). В сенсоре, который мы назвали iATPSnFR 1,0 , линкер L1 был изменен с Thr-Arg на Val-Leu, а L2 с Leu-Gly на Gly-Leu-His. Мы разработали второй датчик (iATPSnFR 1.1 ) с повышенной чувствительностью за счет мутации аминокислот вблизи АТФ-связывающего кармана.iATPSnFR 1,1 отличается от iATPSnFR 1,0 двумя мутациями (Ala95Lys и Ala119Ser; рис.; дополнительный рисунок 1 ). Как iATPSnFR 1,0 , так и iATPSnFR 1,1 демонстрируют заметное улучшение по сравнению с QUEEN-7µ, который не работает как датчик с одной длиной волны, и по сравнению с ATeam для того же диапазона концентраций АТФ (рис.
следующий раздел), но значительно уменьшила вызванные АТФ изменения флуоресценции. Чтобы исправить это, мы повторно оптимизировали L1 и L2 для конструкции cpSFGFP, мутировав аминокислоты в линкерах и слегка изменив их длину; Было проверено около 7000 колоний (рис.). Мы также мутировали аминокислоты (Thr9Val и Asn78Tyr), предсказанные на основе молекулярного моделирования, для уменьшения образования димеров. Благодаря этому процессу мы разработали два датчика, которые демонстрировали значительное АТФ-зависимое увеличение флуоресценции (рис. ). В сенсоре, который мы назвали iATPSnFR 1,0 , линкер L1 был изменен с Thr-Arg на Val-Leu, а L2 с Leu-Gly на Gly-Leu-His. Мы разработали второй датчик (iATPSnFR 1.1 ) с повышенной чувствительностью за счет мутации аминокислот вблизи АТФ-связывающего кармана.iATPSnFR 1,1 отличается от iATPSnFR 1,0 двумя мутациями (Ala95Lys и Ala119Ser; рис.; дополнительный рисунок 1 ). Как iATPSnFR 1,0 , так и iATPSnFR 1,1 демонстрируют заметное улучшение по сравнению с QUEEN-7µ, который не работает как датчик с одной длиной волны, и по сравнению с ATeam для того же диапазона концентраций АТФ (рис. ). Кроме того, вставка cpSFGFP в Queen не приводила к появлению сенсора с усилением флуоресценции, вызванной АТФ. Очищенный iATPSnFR 1.0 имел максимум d F / F ~2,4 и EC 50 ~ 120 мкМ, тогда как очищенный iATPSnFR 1,1 имел максимум d F ~ / F и F . 50 ~50 мкМ (рис. ). Очищенные iATPSnFR не были чувствительны к АДФ, АМФ или аденозину в концентрациях, эквивалентных АТФ (рис. ). Оба белка показали сходные спектры флуоресценции (пик возбуждения 490 нм, пик эмиссии 512 нм; рис. ). В присутствии АТФ наблюдалось увеличение пика возбуждения и эмиссии.На дополнительном рисунке 2 показано, что пик флуоресценции при 400 нм, который наблюдается во многих датчиках на основе GFP, включая QUEEN, был по существу независимым от АТФ. Таким образом, наш скрининг на основе бактерий дал генетически кодируемый сенсор АТФ с микромолярной чувствительностью, большой интенсивностью возбужденного излучения 488 нм d F / F и небольшой чувствительностью к метаболитам АТФ (рис.
). Кроме того, вставка cpSFGFP в Queen не приводила к появлению сенсора с усилением флуоресценции, вызванной АТФ. Очищенный iATPSnFR 1.0 имел максимум d F / F ~2,4 и EC 50 ~ 120 мкМ, тогда как очищенный iATPSnFR 1,1 имел максимум d F ~ / F и F . 50 ~50 мкМ (рис. ). Очищенные iATPSnFR не были чувствительны к АДФ, АМФ или аденозину в концентрациях, эквивалентных АТФ (рис. ). Оба белка показали сходные спектры флуоресценции (пик возбуждения 490 нм, пик эмиссии 512 нм; рис. ). В присутствии АТФ наблюдалось увеличение пика возбуждения и эмиссии.На дополнительном рисунке 2 показано, что пик флуоресценции при 400 нм, который наблюдается во многих датчиках на основе GFP, включая QUEEN, был по существу независимым от АТФ. Таким образом, наш скрининг на основе бактерий дал генетически кодируемый сенсор АТФ с микромолярной чувствительностью, большой интенсивностью возбужденного излучения 488 нм d F / F и небольшой чувствительностью к метаболитам АТФ (рис. ).
).
Тестирование сенсоров АТФ в клетках HEK293
Первоначально мы тестировали сенсоры в клетках HEK293 с помощью конфокальной визуализации.На рисунке показаны три примера iATPSnFR 1,0 и iATPSnFR 1,1 , направленных на клеточную поверхность, временно экспрессированных в клетках HEK293 в течение 24 часов. Изображения показаны до и во время применения АТФ (1 мМ). Соотношение изображений +АТФ и -АТФ также показано справа для каждой серии изображений (d F ). Из этих изображений было ясно, что оба iATPSnFR были экспрессированы в клетках HEK293, нацелены на поверхность клеток, и что оба реагировали заметно заметным увеличением флуоресценции, локализованным на краях клеток (рис.; представитель клеток 290 и 191 для iATPSnFR 1,0 и iATPSnFR 1,1 , соответственно, из четырех отдельных трансфекций каждая). Дополнительные видеоролики 1 и 2 для iATPSnFR 1.0 и iATPSnFR 1.1 также иллюстрируют этот момент. Кривые под каждым набором изображений на рисунке показывают репрезентативные кривые для различных показанных концентраций АТФ, а графики показывают зависимость доза-реакция. Из таких экспериментов мы сделали вывод, что как iATPSnFR 1.0 и iATPSnFR 1.1 функционировали как сенсоры АТФ при внеклеточном отображении на плазматической мембране. Кроме того, в клетках HEK293, совместно трансфицированных цитоплазматическим mCherry и отображаемым на мембране iATPSnFR 1,0 или iATPSnFR 1,1 (рис. ), наблюдалась отчетливая флуоресценция iATPSnFR на краях клеток. Напротив, флуоресценция в цитоплазме в основном отсутствовала, что указывало на правильное перемещение через мембрану (рис. ).
Кривые под каждым набором изображений на рисунке показывают репрезентативные кривые для различных показанных концентраций АТФ, а графики показывают зависимость доза-реакция. Из таких экспериментов мы сделали вывод, что как iATPSnFR 1.0 и iATPSnFR 1.1 функционировали как сенсоры АТФ при внеклеточном отображении на плазматической мембране. Кроме того, в клетках HEK293, совместно трансфицированных цитоплазматическим mCherry и отображаемым на мембране iATPSnFR 1,0 или iATPSnFR 1,1 (рис. ), наблюдалась отчетливая флуоресценция iATPSnFR на краях клеток. Напротив, флуоресценция в цитоплазме в основном отсутствовала, что указывало на правильное перемещение через мембрану (рис. ).
Репрезентативные изображения и кривые для ответов, измеренных с помощью iATPSnFRs клеточной поверхности. a На верхних панелях показаны три примера ответов АТФ для клеток HEK293, экспрессирующих iATPSnFR 1,0 . Нижние панели показывают репрезентативные следы и среднюю кривую доза-реакция для 290 клеток. b Как в ( a ), но для iATPSnFR 1.1 . Кривая доза-реакция представляет собой среднее значение для 191 клетки. На кривых доза-реакция планки погрешностей представляют собой стандартную ошибку среднего. а в некоторых случаях меньше, чем символы, используемые для среднего значения. n Номера представлены на панелях рисунка и относятся к количеству клеток
b Как в ( a ), но для iATPSnFR 1.1 . Кривая доза-реакция представляет собой среднее значение для 191 клетки. На кривых доза-реакция планки погрешностей представляют собой стандартную ошибку среднего. а в некоторых случаях меньше, чем символы, используемые для среднего значения. n Номера представлены на панелях рисунка и относятся к количеству клеток
Характеристика iATPSnFRs клеточной поверхности в клетках HEK293. a Отдельные плоскости из конфокальной z-стеки временно трансфицированных клеток HEK293, экспрессирующих цитоплазматический mCherry и iATPSnFR с отображением на мембране 1,0 . b То же, что и в ( a ), но для iATPSnFR 1,1 временно экспрессируется в клетках HEK293. c Средние кривые для iATPSnFR 1,1 (зеленый) и iATPSnFR 1,0 (красный) в диапазоне концентраций от 10 мкМ до 3 мМ, измеренные с помощью широкопольной эпифлуоресцентной визуализации. d , e Кривые доза-реакция для iATPSnFR 1.0 и iATPSnFR 1.1 , отображаемые на поверхности клеток HEK293. Показаны чувствительность (EC 50 ), общий динамический диапазон (макс.) и кооперативность (нГн). f Репрезентативный Вестерн-блот из клеток HEK293, трансфицированных iATPSnFR 1,0 или iATPSnFR 1,1 , зондированных на наличие GFP (iATPSnFR) и GAPDH (контроль загрузки). Полностью неразрезанный гель показан на дополнительной фигуре 9 . На панели ( f ) график показывает количественную оценку экспрессии iATPSnFR по четырем вестерн-блоттингам, нормализованным по контролю загрузки GAPDH.Круг представляет собой среднее значение, прямоугольник — стандартную ошибку, усы — стандартное отклонение, а горизонтальная линия — медиану. Данные сравнивали с непарным тестом Student t . На других панелях планки погрешностей представляют собой sem. а в некоторых случаях меньше, чем символы, используемые для среднего значения.
d , e Кривые доза-реакция для iATPSnFR 1.0 и iATPSnFR 1.1 , отображаемые на поверхности клеток HEK293. Показаны чувствительность (EC 50 ), общий динамический диапазон (макс.) и кооперативность (нГн). f Репрезентативный Вестерн-блот из клеток HEK293, трансфицированных iATPSnFR 1,0 или iATPSnFR 1,1 , зондированных на наличие GFP (iATPSnFR) и GAPDH (контроль загрузки). Полностью неразрезанный гель показан на дополнительной фигуре 9 . На панели ( f ) график показывает количественную оценку экспрессии iATPSnFR по четырем вестерн-блоттингам, нормализованным по контролю загрузки GAPDH.Круг представляет собой среднее значение, прямоугольник — стандартную ошибку, усы — стандартное отклонение, а горизонтальная линия — медиану. Данные сравнивали с непарным тестом Student t . На других панелях планки погрешностей представляют собой sem. а в некоторых случаях меньше, чем символы, используемые для среднего значения. Номера n представлены на панелях рисунка и относятся к количеству клеток для панелей ( c и d ) и независимым оценкам для панелей ( f )
Номера n представлены на панелях рисунка и относятся к количеству клеток для панелей ( c и d ) и независимым оценкам для панелей ( f )
Без оптических срезов и с использованием широкопольной эпифлуоресцентной визуализации, оба iATPSnFR 1.0 и iATPSnFR 1,1 продемонстрировали зависящее от концентрации увеличение флуоресценции, которое было быстрым, сохранялось в АТФ и возвращалось к исходному уровню после вымывания АТФ (рис. ). iATPSnFR 1,0 показал EC 50 ~ 350 мкМ с максимальным значением d F / F ~ 1,0 (рис. ). Точно так же iATPSnFR 1,1 показал EC 50 ~ 140 мкМ с максимальным значением d F / F ~ 0,9 (рис. ). Таким образом, наблюдалась ~3-кратная потеря чувствительности к АТФ и уменьшение пикового ответа в обоих сенсорах, когда они экспонировались внеклеточно, по сравнению с растворимыми белками (сходные эффекты наблюдаются с другими сенсорами, экспонируемыми на мембране 26 , 27 , 31 ). Во время этих экспериментов мы заметили, что клетки, экспрессирующие iATPSnFR 1,1 , были тусклее, чем клетки, экспрессирующие iATPSnFR 1,0 , и мы обнаружили более низкие, но поддающиеся измерению уровни экспрессии iATPSnFR 1,1 по отношению к iATPSnFR 1,0 (рис. ). Тем не менее, оба были выражены. Так, iATPSnFR 1.1 имеет более низкую ЕС АТФ 50 (более чувствителен), но выражен не так хорошо, как iATPSnFR 1.0 . iATPSnFR 1.1 также продемонстрировал несколько большее внутриклеточное накопление, чем iATPSnFR 1.0 (рис. ).
Во время этих экспериментов мы заметили, что клетки, экспрессирующие iATPSnFR 1,1 , были тусклее, чем клетки, экспрессирующие iATPSnFR 1,0 , и мы обнаружили более низкие, но поддающиеся измерению уровни экспрессии iATPSnFR 1,1 по отношению к iATPSnFR 1,0 (рис. ). Тем не менее, оба были выражены. Так, iATPSnFR 1.1 имеет более низкую ЕС АТФ 50 (более чувствителен), но выражен не так хорошо, как iATPSnFR 1.0 . iATPSnFR 1.1 также продемонстрировал несколько большее внутриклеточное накопление, чем iATPSnFR 1.0 (рис. ).
Кинетика и лиганд-специфичность сенсоров АТФ
Используя быстродействующий переключатель растворов, мы оценили кинетику ответа отображаемых в клетках iATPSnFR 1,0 и iATPSnFR 1,1 по отношению к iGluSnFR, который демонстрирует быстрые скорости включения и выключения 26 , 37 . Постоянные времени увеличения флуоресценции ( τ на ) зависели от дозы и были значительно быстрее для iATPSnFR 1,1 , чем для iATPSnFR 1.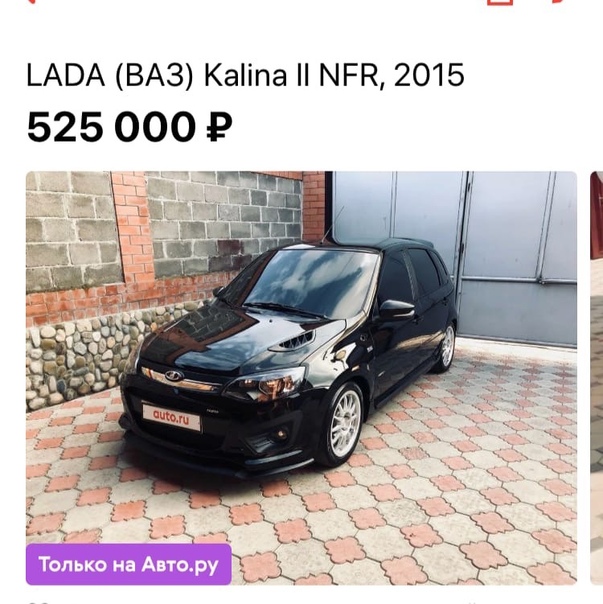 0 (рис. ). Напротив, константы времени для возврата к исходному уровню ( τ от ) АТФ-зависимого увеличения флуоресценции не отличались между двумя конструкциями, что означает, что более высокая равновесная EC 50 iATPSnFR 1,0 была обусловлена из-за более медленной ассоциации АТФ с сенсором или из-за более медленной последующей связи связывания АТФ с изменением флуоресценции. Для сравнения, iGluSnFR был значительно быстрее, чем оба iATPSnFR (рис. ).
0 (рис. ). Напротив, константы времени для возврата к исходному уровню ( τ от ) АТФ-зависимого увеличения флуоресценции не отличались между двумя конструкциями, что означает, что более высокая равновесная EC 50 iATPSnFR 1,0 была обусловлена из-за более медленной ассоциации АТФ с сенсором или из-за более медленной последующей связи связывания АТФ с изменением флуоресценции. Для сравнения, iGluSnFR был значительно быстрее, чем оба iATPSnFR (рис. ).
Оценка кинетики и лигандной селективности сенсоров АТФ на клеточной поверхности. a Следы кинетики iATPSnFR 1,0 , iATPSnFR 1,1 и iGluSnFR из экспериментов по быстрому изменению раствора. Мы могли менять решения менее чем за ~10 мс. b Показывает τ на и τ на для iATPSnFR 1,0 и iATPSnFR 1,1 при различных концентрациях АТФ. c Показывает реакцию iATPSnFR 1,0 и iATPSnFR 1,1 на АТФ, АДФ, АМФ и аденозин (аппликации 1 мМ). d Реакция iATPSnFR 1,0 и iATPSnFR 1,1 на АТФ, ГТФ и УТФ (аппликации 1 мМ). e Следы iATPSnFR 1,0 и iATPSnFR 1,1 с тремя различными концентрациями АТФ (1, 0,3 и 0,1 мМ) с 1 мМ АДФ и без него. АДФ мало влияет на ответы iATPSnFR. Столбики погрешностей представляют собой sem. а в некоторых случаях меньше, чем символы, используемые для среднего значения. Номера n приведены на панелях рисунка и относятся к числу клеток
d Реакция iATPSnFR 1,0 и iATPSnFR 1,1 на АТФ, ГТФ и УТФ (аппликации 1 мМ). e Следы iATPSnFR 1,0 и iATPSnFR 1,1 с тремя различными концентрациями АТФ (1, 0,3 и 0,1 мМ) с 1 мМ АДФ и без него. АДФ мало влияет на ответы iATPSnFR. Столбики погрешностей представляют собой sem. а в некоторых случаях меньше, чем символы, используемые для среднего значения. Номера n приведены на панелях рисунка и относятся к числу клеток
АТФ последовательно расщепляется до АДФ, АМФ и аденозина под действием АТФаз и других АТФ-утилизирующих ферментов в цитозоле и на поверхности клеток.В соответствии с данными бактериального лизата (рис. ), iATPSnFR клеточной поверхности не были чувствительны к АДФ, АМФ или аденозину при концентрации 1 мМ (рис. ). Кроме того, iATPSnFRs не были чувствительны к 1 мМ GTP или 1 мМ UTP (рис. ), другим нуклеозидтрифосфатам, обнаруженным в цитозоле и во внеклеточном пространстве. Поскольку ожидается, что внеклеточное пространство будет содержать смесь АТФ и АДФ, мы оценили, влияет ли 1 мМ АДФ на реакцию на 0,1, 0,3 и 1 мМ АТФ для iATPSnFR 1,0 и iATPSnFR 1.1 . Это не так (рис. ). Таким образом, iATPSnFR воспринимают АТФ с незначительной реакцией на продукты деградации или на общие конкурирующие нуклеотиды и нуклеозиды.
Поскольку ожидается, что внеклеточное пространство будет содержать смесь АТФ и АДФ, мы оценили, влияет ли 1 мМ АДФ на реакцию на 0,1, 0,3 и 1 мМ АТФ для iATPSnFR 1,0 и iATPSnFR 1.1 . Это не так (рис. ). Таким образом, iATPSnFR воспринимают АТФ с незначительной реакцией на продукты деградации или на общие конкурирующие нуклеотиды и нуклеозиды.
Экспрессия iATPSnFR
1,0 в астроцитах и нейронах Для характеристики астроцитов и нейронов мы сосредоточились на iATPSnFR 1,0 , учитывая его лучшую экспрессию. Мы совместно экспрессировали iATPSnFR 1,0 с mCherry в культивируемой астроглии U373MG и наблюдали хороший перенос на клеточную поверхность (рис. ), хотя также имелся внутриклеточный пул сенсора.Мы подтвердили, что iATPSnFR 1,0 реагировал на применение 300 мкМ АТФ увеличением флуоресценции (рис. ). Аналогичные данные были получены при экспрессии iATPSnFR 1.0 в первичных культурах нейронов гиппокампа крысы (рис. ). Для астроглии U373MG мы определили, что iATPSnFR 1,0 EC 50 составляет 400 мкМ с максимальным значением d F / F , равным 1,1 (рис. ), аналогично данным по клеткам HEK293. В нейронах гиппокампа iATPSnFR 1,0 правильно локализовался в мембранных отростках, а применение ~ 1 мМ АТФ вызывало усиление флуоресценции.В нейронах гиппокампа АТФ EC 50 составляло 630 мкМ, а максимальное значение d F / F составляло ~ 1,5 (рис. ). Таким образом, наши опыты с iATPSnFR 1,0 в растворе, в клетках HEK293, в астроглии и в нейронах гиппокампа демонстрируют, что ослабление связывания АТФ и уменьшение d F / F в результате привязки сенсора к внешней стороне бислой мембраны не зависит от клетки-хозяина. Мы наблюдали изменение чувствительности у других сенсоров на мембране 26 , 27 , 31 ; мы предполагаем, что это происходит из-за стерических ограничений головных групп липидов и мембранных белков.
). Для астроглии U373MG мы определили, что iATPSnFR 1,0 EC 50 составляет 400 мкМ с максимальным значением d F / F , равным 1,1 (рис. ), аналогично данным по клеткам HEK293. В нейронах гиппокампа iATPSnFR 1,0 правильно локализовался в мембранных отростках, а применение ~ 1 мМ АТФ вызывало усиление флуоресценции.В нейронах гиппокампа АТФ EC 50 составляло 630 мкМ, а максимальное значение d F / F составляло ~ 1,5 (рис. ). Таким образом, наши опыты с iATPSnFR 1,0 в растворе, в клетках HEK293, в астроглии и в нейронах гиппокампа демонстрируют, что ослабление связывания АТФ и уменьшение d F / F в результате привязки сенсора к внешней стороне бислой мембраны не зависит от клетки-хозяина. Мы наблюдали изменение чувствительности у других сенсоров на мембране 26 , 27 , 31 ; мы предполагаем, что это происходит из-за стерических ограничений головных групп липидов и мембранных белков.
Характеристика клеточной поверхности и цитозоля iATPSnFR 1,0 в астроцитах и нейронах. a Одна плоскость из конфокального z-стека клеток U373MG, экспрессирующих цитоплазматическую mCherry и отображающую на мембране iATPSnFR 1,0 . b Конфокальные изображения клеток U373MG, экспрессирующих iATPSnFR 1,0 , до применения АТФ (контроль) и во время применения 300 мкМ АТФ (+АТФ), а также изменение флуоресценции (d F ). c Конфокальные изображения нейронов гиппокампа до аппликации АТФ (контроль) и во время аппликации 1 мМ АТФ, а также изменение флуоресценции (d F ). d , e Кривые доза-реакция для iATPSnFR 1,0 при отображении на поверхности нейронов астроглии U373MG и гиппокампа. f Конфокальное изображение клетки астроглии U373MG, трансфицированной iATPSnFR 1,0 , до введения 3 мМ АТФ (контроль), во время воздействия (АТФ), а также изменение флуоресценции (d F ). Средний след от всех клеток с с.э.м. также показано. г Репрезентативный астроцит гиппокампа в остром срезе головного мозга, экспрессирующий iATPSnFR 1.0 до применения АТФ (контроль), во время применения 3 мМ АТФ (АТФ) и dF. Средний след от всех клеток с с.э.м. также показано. ч Сводка диаграммы рассеяния эффекта 3 мМ АТФ на iATPSnFR 1,0 выражена в указанных типах клеток. На панели ( h ) кружок представляет среднее значение, прямоугольник — стандартную ошибку, усы — стандартное отклонение, а горизонтальная линия — медиану. p Значения показаны для статистического сравнения с контрольным буферным раствором с использованием непарных тестов Стьюдента t .Сокращения: гиппокамп тазобедренного сустава, SC спинной мозг, MSNs средние шипиковые нейроны. Обратите внимание на сломанные оси и . На других панелях планки погрешностей представляют собой sem. а в некоторых случаях меньше, чем символы, используемые для среднего значения.
Средний след от всех клеток с с.э.м. также показано. г Репрезентативный астроцит гиппокампа в остром срезе головного мозга, экспрессирующий iATPSnFR 1.0 до применения АТФ (контроль), во время применения 3 мМ АТФ (АТФ) и dF. Средний след от всех клеток с с.э.м. также показано. ч Сводка диаграммы рассеяния эффекта 3 мМ АТФ на iATPSnFR 1,0 выражена в указанных типах клеток. На панели ( h ) кружок представляет среднее значение, прямоугольник — стандартную ошибку, усы — стандартное отклонение, а горизонтальная линия — медиану. p Значения показаны для статистического сравнения с контрольным буферным раствором с использованием непарных тестов Стьюдента t .Сокращения: гиппокамп тазобедренного сустава, SC спинной мозг, MSNs средние шипиковые нейроны. Обратите внимание на сломанные оси и . На других панелях планки погрешностей представляют собой sem. а в некоторых случаях меньше, чем символы, используемые для среднего значения. n числа указаны на панелях рисунка и относятся к количеству клеток D промоутер 38 .После микроинъекций in vivo в гиппокамп iATPSnFR 1,0 совместно локализовался с S100β-положительными астроцитами, но не с NeuN-позитивными нейронами (дополнительная фигура 3 ). Нанесение АТФ в ванну с многоклеточными препаратами, такими как срезы головного мозга, сопряжено с проблемами, вызванными быстрым гидролизом АТФ эктоАТФазами клеточной поверхности. Чтобы свести к минимуму такие эффекты, мы вдували АТФ через стеклянную пипетку с пикосприцером. Напыление 3 мМ АТФ (в течение 5 с) на астроглию U373MG привело к ~ 0.7 d F / F (рис. ). Подобные, хотя и меньшие, ответы наблюдались, когда мы наносили АТФ на астроциты, экспрессирующие iATPSnFR 1,0 , в срезах гиппокампа и спинного мозга или на средние шипиковые нейроны D1 или D2, экспрессирующие iATPSnFR, в стриатуме (рис. ). В совокупности эти данные показывают, что iATPSnFR может экспрессироваться и надежно реагировать на поверхности клеток после экспрессии in vivo.
n числа указаны на панелях рисунка и относятся к количеству клеток D промоутер 38 .После микроинъекций in vivo в гиппокамп iATPSnFR 1,0 совместно локализовался с S100β-положительными астроцитами, но не с NeuN-позитивными нейронами (дополнительная фигура 3 ). Нанесение АТФ в ванну с многоклеточными препаратами, такими как срезы головного мозга, сопряжено с проблемами, вызванными быстрым гидролизом АТФ эктоАТФазами клеточной поверхности. Чтобы свести к минимуму такие эффекты, мы вдували АТФ через стеклянную пипетку с пикосприцером. Напыление 3 мМ АТФ (в течение 5 с) на астроглию U373MG привело к ~ 0.7 d F / F (рис. ). Подобные, хотя и меньшие, ответы наблюдались, когда мы наносили АТФ на астроциты, экспрессирующие iATPSnFR 1,0 , в срезах гиппокампа и спинного мозга или на средние шипиковые нейроны D1 или D2, экспрессирующие iATPSnFR, в стриатуме (рис. ). В совокупности эти данные показывают, что iATPSnFR может экспрессироваться и надежно реагировать на поверхности клеток после экспрессии in vivo.
Затем мы исследовали использование цитозольного iATPSnFR 1,0 для отслеживания внутриклеточного АТФ, который колеблется от 1 до 3 мМ в клетках млекопитающих 39 .Мы экспрессировали цитозольный iATPSnFR 1,0 в астроглии U373MG и обнаружили значительное снижение внутриклеточной флуоресценции при переходе на 0 мМ глюкозы (дополнительный рисунок 4a ) или при применении 10 мМ 2-дезоксиглюкозы (2DOG; дополнительный рисунок 4b 900). ингибитор гликолиза 40 . Точно так же после экспрессии in vivo с AAV мы измерили значительное снижение уровней внутриклеточного АТФ, о чем свидетельствует флуоресценция iATPSnFR 1.0 в астроцитах гиппокампа в 0 мМ глюкозы (дополнительная фигура 4c ; флуоресценция коэкспрессированного tdTomato не изменилась, что свидетельствует об изменении формы и объема клеток или других неспецифических артефактах в качестве причины). Мы повторили эти эксперименты для астроцитов гиппокампа и пирамидных нейронов CA1, как в соматах, так и вдоль отростков (дополнительная фигура 4d ): кислородно-глюкозная депривация (OGD) вызывала значительное снижение внутриклеточного уровня АТФ. Мы обнаружили, что снижение флуоресценции астроцитов было вдвое больше, чем у нейронов (дополнительный рисунок).Такие ответы были полностью обратимы для астроцитов, но лишь частично обратимы для нейронов (дополнительная фигура 4d ). Кроме того, для начала снижения уровня АТФ в астроцитах потребовалось больше времени (~ 10 минут в OGD). Однако, как только АТФ начал уменьшаться в астроцитах, потребовалось 8,0 ± 0,5 мин ( n = 37), чтобы уменьшиться на 50%, тогда как в соматах и дендритах пирамидных нейронов СА1 это заняло 13,5 ± 0,8 мин ( n 2)= 6 и 17,4 ± 0,9 мин ( n = 27) соответственно ( p < 0.0001 по ANOVA и апостериорным тестам Даннета, здесь ± указывает плюс/минус стандартная ошибка среднего). Основываясь на титровании на рис. , мы оцениваем, что уровни АТФ упали на ~ 0,5 мМ с> 1 мМ. Таким образом, во время депривации глюкозы уровень АТФ в астроцитах быстро падает, но полностью восстанавливается при восстановлении уровня глюкозы.
Мы обнаружили, что снижение флуоресценции астроцитов было вдвое больше, чем у нейронов (дополнительный рисунок).Такие ответы были полностью обратимы для астроцитов, но лишь частично обратимы для нейронов (дополнительная фигура 4d ). Кроме того, для начала снижения уровня АТФ в астроцитах потребовалось больше времени (~ 10 минут в OGD). Однако, как только АТФ начал уменьшаться в астроцитах, потребовалось 8,0 ± 0,5 мин ( n = 37), чтобы уменьшиться на 50%, тогда как в соматах и дендритах пирамидных нейронов СА1 это заняло 13,5 ± 0,8 мин ( n 2)= 6 и 17,4 ± 0,9 мин ( n = 27) соответственно ( p < 0.0001 по ANOVA и апостериорным тестам Даннета, здесь ± указывает плюс/минус стандартная ошибка среднего). Основываясь на титровании на рис. , мы оцениваем, что уровни АТФ упали на ~ 0,5 мМ с> 1 мМ. Таким образом, во время депривации глюкозы уровень АТФ в астроцитах быстро падает, но полностью восстанавливается при восстановлении уровня глюкозы. Напротив, уровни АТФ в нейронах падают медленно и не могут восстановиться до нормального уровня в тех же условиях.
Напротив, уровни АТФ в нейронах падают медленно и не могут восстановиться до нормального уровня в тех же условиях.
mRuby-iATPSnFR
1.0 в качестве ратиометрического сенсора АТФ Мы слили красный флуоресцентный белок mRuby 41 с N-концом iATPSnFR 1.0 для создания логометрического датчика (рис.; mRuby-iATPSnFR 1.0 ). Конструкция mRuby-iATPSnFR 1.0 была идентична конструкции iATPSnFR 1.0 по своей чувствительности и вызываемой АТФ d F / F (рис. ). В клетках HEK293 mRuby-iATPSnFR 1,0 демонстрировал явные изменения зеленой флуоресценции в присутствии 2DOG, тогда как красная флуоресценция mRuby не менялась (рис. ; дополнительная фигура 5 ). Обратите внимание, что на дополнительном рисунке 5 показано поле зрения, но анализы проводились на отдельных клетках, а не путем усреднения по всем полям зрения.Точно так же после экспрессии AAV in vivo мы измерили значительное снижение внутриклеточного АТФ, о чем свидетельствует флуоресценция iATPSnFR 1,0 в астроцитах гиппокампа и соматах и дендритах пирамидных нейронов при переходе на растворы, имитирующие ишемию (т. е. OGD), тогда как в mRuby изменений не было (рис.; дополнительная фигура 6 ). Интересно, что уровни АТФ возвращались к нормальному уровню при повторном применении физиологических буферов в астроцитах, но оставались низкими в соматах и дендритах нейронов (рис.). Мы также провели два необходимых контрольных эксперимента. Во-первых, мы проверили, реагировал ли сам iATPSnFR 1.0 на 2DOG снижением флуоресценции. Это было не так (дополнительная фигура 7 ). Во-вторых, мы измерили внутриклеточные изменения pH в клетках HEK293 во время применения 2DOG. Используя конфокальную микроскопию (Ex 473 нм, Em 525 нм), мы обнаружили тонкое 10% снижение флуоресценции индикатора pH BCECF, что предполагает подкисление внутриклеточного компартмента на ~0.2 единицы рН. Однако это подкисление заняло ~ 45 минут, чтобы достичь значимости в присутствии 2DOG, и не было обратимым при повторном применении глюкозы (дополнительная фигура 7b ). Для дальнейшего изучения изменений pH мы также использовали эпифлуоресцентную логометрическую визуализацию с двумя длинами волн с помощью BCECF-AM (Ex 505 и 440 нм, Em 535 нм) и обнаружили, что соотношение 505/440 уменьшилось с 1,3 ± 0,03 до 1,2 ± 0,03 через 20 минут.
е. OGD), тогда как в mRuby изменений не было (рис.; дополнительная фигура 6 ). Интересно, что уровни АТФ возвращались к нормальному уровню при повторном применении физиологических буферов в астроцитах, но оставались низкими в соматах и дендритах нейронов (рис.). Мы также провели два необходимых контрольных эксперимента. Во-первых, мы проверили, реагировал ли сам iATPSnFR 1.0 на 2DOG снижением флуоресценции. Это было не так (дополнительная фигура 7 ). Во-вторых, мы измерили внутриклеточные изменения pH в клетках HEK293 во время применения 2DOG. Используя конфокальную микроскопию (Ex 473 нм, Em 525 нм), мы обнаружили тонкое 10% снижение флуоресценции индикатора pH BCECF, что предполагает подкисление внутриклеточного компартмента на ~0.2 единицы рН. Однако это подкисление заняло ~ 45 минут, чтобы достичь значимости в присутствии 2DOG, и не было обратимым при повторном применении глюкозы (дополнительная фигура 7b ). Для дальнейшего изучения изменений pH мы также использовали эпифлуоресцентную логометрическую визуализацию с двумя длинами волн с помощью BCECF-AM (Ex 505 и 440 нм, Em 535 нм) и обнаружили, что соотношение 505/440 уменьшилось с 1,3 ± 0,03 до 1,2 ± 0,03 через 20 минут. 2DOG, но не восстановился при повторном применении глюкозы (1,1 ± 0,02) в течение 20 мин ( n = 144).Следовательно, в соответствии с предыдущей работой с использованием 2DOG 20 мы измерили переменное и незначительное подкисление в клетках HEK293 с применением 2DOG, но эти изменения не могли объяснить падение флуоресценции mRuby-iATPSnFR 1,0 , которое мы связываем с падением в концентрации АТФ, о которой сообщалось ранее 20 . Например, из калибровочных экспериментов, представленных на рис. (см. следующий раздел), ожидается, что подкисление с pH 7,4 до 7,2 вызовет снижение флуоресценции АТФ-связанного mRuby-iATPSnFR 1 на 9%.0 , тогда как с приложениями 2DOG мы измерили изменение от 40 до 50% (рис. ). Таким образом, в соответствии с прошлыми исследованиями 20 такие предполагаемые изменения АТФ были более значительными, более быстрыми и обратимыми в клетках HEK293 (рис. ). Мы не могли использовать BCECF в качестве индикатора pH для экспериментов со срезами мозга, потому что наша предыдущая работа показала, что органические красители версии ацетоксиметилового эфира (AM), проникающие через мембрану, плохо/ненадежно поглощаются срезами мозга взрослых 38 , 42 .
2DOG, но не восстановился при повторном применении глюкозы (1,1 ± 0,02) в течение 20 мин ( n = 144).Следовательно, в соответствии с предыдущей работой с использованием 2DOG 20 мы измерили переменное и незначительное подкисление в клетках HEK293 с применением 2DOG, но эти изменения не могли объяснить падение флуоресценции mRuby-iATPSnFR 1,0 , которое мы связываем с падением в концентрации АТФ, о которой сообщалось ранее 20 . Например, из калибровочных экспериментов, представленных на рис. (см. следующий раздел), ожидается, что подкисление с pH 7,4 до 7,2 вызовет снижение флуоресценции АТФ-связанного mRuby-iATPSnFR 1 на 9%.0 , тогда как с приложениями 2DOG мы измерили изменение от 40 до 50% (рис. ). Таким образом, в соответствии с прошлыми исследованиями 20 такие предполагаемые изменения АТФ были более значительными, более быстрыми и обратимыми в клетках HEK293 (рис. ). Мы не могли использовать BCECF в качестве индикатора pH для экспериментов со срезами мозга, потому что наша предыдущая работа показала, что органические красители версии ацетоксиметилового эфира (AM), проникающие через мембрану, плохо/ненадежно поглощаются срезами мозга взрослых 38 , 42 . В целом, эксперименты mRuby-iATPSnFR 1,0 позволяют предположить, что этот датчик можно использовать в качестве логометрического датчика клеточно-специфических внутриклеточных уровней АТФ, с той оговоркой, что параллельные эксперименты для оценки изменений pH необходимо проводить в каждом конкретном случае. всякий раз, когда используется mRuby-iATPSnFR 1.0 . Затем мы непосредственно оценили чувствительность сенсоров к pH.
В целом, эксперименты mRuby-iATPSnFR 1,0 позволяют предположить, что этот датчик можно использовать в качестве логометрического датчика клеточно-специфических внутриклеточных уровней АТФ, с той оговоркой, что параллельные эксперименты для оценки изменений pH необходимо проводить в каждом конкретном случае. всякий раз, когда используется mRuby-iATPSnFR 1.0 . Затем мы непосредственно оценили чувствительность сенсоров к pH.
Характеристика цитозольного mRuby-iATPSnFR 1,0 . a Мультипликационная схема mRuby-iATPSnFR 1.0 : Связывание АТФ схематизировано, чтобы показать увеличение флуоресценции cpSFGFP, тогда как флуоресценция mRuby остается неизменной. b Кривые доза-реакция для iATPSnFR 1,0 по отношению к mRuby-iATPSnFR 1,0 в клеточных лизатах HEK293. c Изменения флуоресценции mRuby-iATPSnFR 1,0 (зеленый и красный каналы) до, во время и после ингибирования гликолиза 2DOG в клетках HEK293. d Изменения флуоресценции mRuby-iATPSnFR 1.0 (зеленый и красный каналы) во время кислородно-глюкозной депривации в срезах гиппокампа головного мозга. Показаны данные по астроцитам, а также телам и дендритам пирамидных нейронов СА1. Столбики погрешностей представляют собой sem. а в некоторых случаях меньше, чем символы, используемые для среднего значения. n Номера приведены на панелях рисунка и относятся к количеству клеток
d Изменения флуоресценции mRuby-iATPSnFR 1.0 (зеленый и красный каналы) во время кислородно-глюкозной депривации в срезах гиппокампа головного мозга. Показаны данные по астроцитам, а также телам и дендритам пирамидных нейронов СА1. Столбики погрешностей представляют собой sem. а в некоторых случаях меньше, чем символы, используемые для среднего значения. n Номера приведены на панелях рисунка и относятся к количеству клеток
Оценки влияния pH на iATPSnFR 1,0 . a–d Влияние рН на несвязанную и связанную с АТФ флуоресценцию указанных конструкций. e , f Сравнение влияния АТФ на флуоресценцию указанных конструкций при рН 6,4 и 7,4. Столбики погрешностей представляют собой sem. а в некоторых случаях меньше, чем символы, используемые для среднего значения. n числа представлены на панелях рисунков и относятся к количеству клеток в панели ( a ) и количеству независимых оценок в ( b – f )
pH-чувствительность iATPSnFR
1,0 и mRuby-iATPSnFR 1. 0
0 Как и для всех сенсоров на основе GFP 43 , ожидается, что флуоресценция iATPSnFRs будет чувствительна к pH с зависимостью H + , в значительной степени определяемой GFP. Мы провели определенный набор экспериментов, чтобы оценить это как руководство для будущего использования датчиков. Мы сосредоточились на iATPSnFR 1.0 , потому что он использовался для большей части работы, представленной в этом исследовании. Во-первых, мы измерили флуоресценцию внеклеточно представленного iATPSnFR 1,0 на поверхности клеток HEK293 в диапазоне значений pH от 4 до 9 в отсутствие и в присутствии АТФ (рис.). Как и ожидалось, подкисление сильно уменьшило флуоресценцию связанного и несвязанного состояний АТФ. Между pH 6,5 и 9 наблюдалось сильное увеличение флуоресценции при связывании АТФ. Во-вторых, мы повторили описанные выше эксперименты с иАТПСнФР 1.0 в растворе (из бактериальных лизатов). Мы обнаружили тот же эффект при подкислении для связанного и несвязанного состояний АТФ (рис. ). В-третьих, мы провели аналогичные эксперименты для сенсора mRuby-iATPSnFR 1,0 в растворе (из бактериальных лизатов) при оценке эмиссии флуоресценции в зеленом канале сенсора АТФ и в эталонном красном канале mRuby (рис.). Для сенсоров АТФ мы снова обнаружили, что подкисление снижает флуоресценцию связанных и несвязанных состояний АТФ. Влияние рН mRuby не зависело от АТФ, что указывает на то, что mRuby-iATPSnFR 1,0 можно использовать в качестве логометрического датчика (рис. ). В-четвертых, мы построили кривые доза-реакция АТФ для iATPSnFR 1,0 и mRuby-iATPSnFR 1,0 в растворе при pH 6,4 и 7,4 (рис. ). Эти данные показали, что подкисление снижает реакцию флуоресценции при всех концентрациях АТФ, которые были эффективны для усиления флуоресценции.В целом, эти данные показывают, что iATPSnFR 1.0 и mRuby-iATPSnFR 1.0 функционируют как одноволновые и логометрические датчики АТФ, но действительно проявляют чувствительность к кислому pH, как и ожидалось по природе GFP 43 .
). В-третьих, мы провели аналогичные эксперименты для сенсора mRuby-iATPSnFR 1,0 в растворе (из бактериальных лизатов) при оценке эмиссии флуоресценции в зеленом канале сенсора АТФ и в эталонном красном канале mRuby (рис.). Для сенсоров АТФ мы снова обнаружили, что подкисление снижает флуоресценцию связанных и несвязанных состояний АТФ. Влияние рН mRuby не зависело от АТФ, что указывает на то, что mRuby-iATPSnFR 1,0 можно использовать в качестве логометрического датчика (рис. ). В-четвертых, мы построили кривые доза-реакция АТФ для iATPSnFR 1,0 и mRuby-iATPSnFR 1,0 в растворе при pH 6,4 и 7,4 (рис. ). Эти данные показали, что подкисление снижает реакцию флуоресценции при всех концентрациях АТФ, которые были эффективны для усиления флуоресценции.В целом, эти данные показывают, что iATPSnFR 1.0 и mRuby-iATPSnFR 1.0 функционируют как одноволновые и логометрические датчики АТФ, но действительно проявляют чувствительность к кислому pH, как и ожидалось по природе GFP 43 . Возможно, более полезным является то, что калибровочные кривые, показанные на рис. , представляют собой показатель, по которому можно судить о последствиях подкисления в будущих экспериментах, в которых используются датчики. Сенсоры, вероятно, будут иметь ограниченное применение в средах, где рН более кислый, чем ~6.2. Наконец, мы оценили кинетику связывания АТФ с iATPSnFR 1,0 в растворе при pH 7,4 и обнаружили, что они не следовали типичному поведению псевдопервого порядка (дополнительная фигура 8 ), а вместо этого соответствовали механизму в в которых возникают как более высокие, так и более низкие флуоресцентные состояния (потенциально возникающие в результате мономерных и димерных состояний). Эту возможность можно с пользой изучить в последующих исследованиях.
Возможно, более полезным является то, что калибровочные кривые, показанные на рис. , представляют собой показатель, по которому можно судить о последствиях подкисления в будущих экспериментах, в которых используются датчики. Сенсоры, вероятно, будут иметь ограниченное применение в средах, где рН более кислый, чем ~6.2. Наконец, мы оценили кинетику связывания АТФ с iATPSnFR 1,0 в растворе при pH 7,4 и обнаружили, что они не следовали типичному поведению псевдопервого порядка (дополнительная фигура 8 ), а вместо этого соответствовали механизму в в которых возникают как более высокие, так и более низкие флуоресцентные состояния (потенциально возникающие в результате мономерных и димерных состояний). Эту возможность можно с пользой изучить в последующих исследованиях.
Обсуждение
В настоящем исследовании мы использовали яркий, стабильный каркас cpSFGFP, рациональный дизайн и скрининг библиотеки для выбора датчиков АТФ с желаемыми характеристиками. Мы охарактеризовали датчики АТФ для мониторинга клеточной поверхности и уровней цитозольного АТФ с превосходной специфичностью. Текущие датчики могут быть полезны для изучения уровней цитозольного АТФ и сигналов, связанных с рецепторами P2X7, которые реагируют на АТФ в диапазоне от сотен микромолярных до миллимолярных 44 , но они кажутся непригодными для измерения высвобождения АТФ из астроцитов, которое, вероятно, диапазон сотен наномолярных до микромолярных 45 . Датчики также могут быть полезны в диагностике для определения высоких уровней АТФ в опухолях 46 .Стратегии дальнейшего уточнения рассматриваются в следующих разделах.
Мы свели в таблицу свойства iATPSnFR по отношению к свойствам существующих сенсоров АТФ на основе флуоресценции в дополнительной таблице 1 . Уровни внутриклеточного АТФ обычно находятся в миллимолярном диапазоне и изменяются при патологии. Уровни внеклеточного АТФ обычно низкие в физиологических условиях из-за эктоАТФаз, но несколько десятилетий физиологических исследований и существование рецепторов АТФ на клеточной поверхности с чувствительностью к АТФ между наномолярными и миллимолярными указывает на то, что уровни внеклеточного АТФ также увеличиваются во время определенных процессов, таких как пуринергическая передача 5 , 47 – 52 . Принимая во внимание эти факты, кажется невозможным, чтобы один датчик подходил для мониторинга внутриклеточных и внеклеточных уровней АТФ в диапазоне концентраций, охватывающем 5–6 порядков. Таким образом, датчики АТФ должны быть разработаны для конкретных приложений из доступных (дополнительная таблица 1 ) и описанных здесь.
Принимая во внимание эти факты, кажется невозможным, чтобы один датчик подходил для мониторинга внутриклеточных и внеклеточных уровней АТФ в диапазоне концентраций, охватывающем 5–6 порядков. Таким образом, датчики АТФ должны быть разработаны для конкретных приложений из доступных (дополнительная таблица 1 ) и описанных здесь.
Очевидно, что для лучшего обнаружения небольших колебаний АТФ у живых животных необходимо улучшить чувствительность, d F / F и кинетику иАТФСнФР, что может быть достигнуто с помощью стратегий направленной эволюции с применением иАТФСнФР 1.0 эшафот нам сообщили. Важные мишени для такого мутагенеза включают линкерные области и сам АТФ-связывающий модуль. Кроме того, можно использовать тот же каркас для разработки сенсоров для других нуклеотидов и нуклеозидов, как при разработке сенсоров NADPH, например, 53 . Несколько таких стратегий находятся в стадии реализации. В этом отношении прогресс в создании других датчиков, таких как датчики для Ca 2+ , глутамата и напряжения, освещает: все наилучшие доступные индикаторы были созданы через несколько лет и итеративно после того, как была сообщена полезная отправная точка и стратегия. Вполне возможно, что разработка датчиков АТФ пойдет по тому же пути на основе каркаса iATPSnFR 1.0 или других. Параллельно, однако, АТФ-рецепторы, связанные с G-белком (GPCR), которые демонстрируют диапазон сродства к АТФ и лигандной селективности 47 , потенциально могут быть использованы для разработки сенсоров АТФ, о чем свидетельствуют недавние впечатляющие успехи, достигнутые с образованием ацетилхолина. и датчики дофамина на основе родственных им GPCR 28 – 30 .При оценке этих подходов возможно, что датчики на основе GPCR могут образовывать гетеромеры , , 54, , с эндогенными GPCR, тогда как для датчиков, использующих инженерные каркасы, такие как iATPSnFR, такая возможность менее вероятна. Наконец, датчики АТФ на основе GFP, вероятно, найдут много применений для изучения динамики АТФ, и их следует рассматривать как дополнение к использованию подходов к визуализации АТФ на основе люциферазы, которые, как было показано, работают для внутри- и внеклеточного АТФ в клетках и в клетках.
Вполне возможно, что разработка датчиков АТФ пойдет по тому же пути на основе каркаса iATPSnFR 1.0 или других. Параллельно, однако, АТФ-рецепторы, связанные с G-белком (GPCR), которые демонстрируют диапазон сродства к АТФ и лигандной селективности 47 , потенциально могут быть использованы для разработки сенсоров АТФ, о чем свидетельствуют недавние впечатляющие успехи, достигнутые с образованием ацетилхолина. и датчики дофамина на основе родственных им GPCR 28 – 30 .При оценке этих подходов возможно, что датчики на основе GPCR могут образовывать гетеромеры , , 54, , с эндогенными GPCR, тогда как для датчиков, использующих инженерные каркасы, такие как iATPSnFR, такая возможность менее вероятна. Наконец, датчики АТФ на основе GFP, вероятно, найдут много применений для изучения динамики АТФ, и их следует рассматривать как дополнение к использованию подходов к визуализации АТФ на основе люциферазы, которые, как было показано, работают для внутри- и внеклеточного АТФ в клетках и в клетках. vivo 16 – 19 , 32 .Например, кажется маловероятным, что датчики АТФ на основе флуоресценции могут быть использованы для визуализации целых грызунов, что возможно для датчиков на основе люциферазы 19 . Следовательно, выбор используемого датчика будет во многом зависеть от конкретного решаемого вопроса, и в каждом конкретном случае необходимо будет проводить тщательную оценку вклада pH в сигналы.
vivo 16 – 19 , 32 .Например, кажется маловероятным, что датчики АТФ на основе флуоресценции могут быть использованы для визуализации целых грызунов, что возможно для датчиков на основе люциферазы 19 . Следовательно, выбор используемого датчика будет во многом зависеть от конкретного решаемого вопроса, и в каждом конкретном случае необходимо будет проводить тщательную оценку вклада pH в сигналы.
В целом, описанные здесь iATPSnFR и будущие разработки, которые они позволяют (для адаптации чувствительности и кинетики к конкретным приложениям), предвещают прямую визуализацию пуринергической передачи сигналов 5 по всему телу трансгенных животных.Наши конструкции доступны (дополнительная таблица 2 ), чтобы позволить начать такие исследования параллельно с существующими подходами, обсуждаемыми в предыдущих разделах.
Методы
Клонирование
Кодирующая последовательность QUEEN-7µ была приобретена в виде фрагмента гена gBlock у Integrated DNA Technologies с сайтом рестрикции BglII , предшествующим кодирующей последовательности QUEEN-7µ, и сайтом рестрикции PstI после кодирования последовательность. Этот gBlock был клонирован в собственный модифицированный вектор pRSET-A (ThermoFisher: V35120), названный pHHM.pHHM содержал N-концевую метку His6Gly, используемую для очистки, за которой следовала метка эпитопа гемагглютинина (HA) и сайт множественного клонирования перед интересующим геном, а также метка myc на С-конце гена. Этот вектор использовали для скрининга, потому что он имеет метки HA и myc, присутствующие в pDisplay (ThermoFisher: V66020), векторе, который мы планировали использовать для поверхностной экспрессии в клетках млекопитающих 26 , 55 . Клонирование из pHHM в pDisplay или в pDisplayMini, собственный вектор, в котором отсутствует N-концевой эпитоп HA, присутствующий в pDisplay, было достигнуто с помощью рестрикции плазмид iATPSnFR и pDisplay с BglII и PstI .Клонирование в pZAC-AAV- GfaABC 1 D проводили с использованием ПЦР-амплификации для добавления сайта рестрикции NheI к N-концу iATPSnFR и сайта рестрикции NotI in к С-концу.
Этот gBlock был клонирован в собственный модифицированный вектор pRSET-A (ThermoFisher: V35120), названный pHHM.pHHM содержал N-концевую метку His6Gly, используемую для очистки, за которой следовала метка эпитопа гемагглютинина (HA) и сайт множественного клонирования перед интересующим геном, а также метка myc на С-конце гена. Этот вектор использовали для скрининга, потому что он имеет метки HA и myc, присутствующие в pDisplay (ThermoFisher: V66020), векторе, который мы планировали использовать для поверхностной экспрессии в клетках млекопитающих 26 , 55 . Клонирование из pHHM в pDisplay или в pDisplayMini, собственный вектор, в котором отсутствует N-концевой эпитоп HA, присутствующий в pDisplay, было достигнуто с помощью рестрикции плазмид iATPSnFR и pDisplay с BglII и PstI .Клонирование в pZAC-AAV- GfaABC 1 D проводили с использованием ПЦР-амплификации для добавления сайта рестрикции NheI к N-концу iATPSnFR и сайта рестрикции NotI in к С-концу. кодирующей последовательности iATPSnFR. Клонирование выполняли с использованием обычных методов субклонирования. Обратите внимание, что mRuby, который мы использовали, был mRuby3 минус MDELYK с C-конца.
кодирующей последовательности iATPSnFR. Клонирование выполняли с использованием обычных методов субклонирования. Обратите внимание, что mRuby, который мы использовали, был mRuby3 минус MDELYK с C-конца.
Экспрессия и очистка белков
Векторы, содержащие варианты iATPSnFR, трансформировали в клетки Escherichia coli BL21(DE3) (без pLysS).Белки экспрессировали путем выращивания E. coli в жидких средах для аутоиндукции 56 с добавлением 100 мкг/мл ампициллина при 30°C в течение не менее 18 часов. После экспрессии бактерии собирали при 1500 г в течение 20 мин. Осадок ресуспендировали в PBS с 1М NaCl и замораживали. Белок экстрагировали с помощью лизиса замораживанием-оттаиванием и обработки ультразвуком. После оттаивания к раствору перед обработкой ультразвуком добавляли 1 мкМ PMSF. После обработки ультразвуком сырой лизат сначала осветляли при 4000 г в течение 20 мин.Затем супернатант выделяли и дополнительно очищали центрифугированием при 35 000 g в течение 45 мин. Белки очищали с помощью аффинной хроматографии на Ni-NTA-агарозе (колонка 5 мл). Белки iATPSnFR элюировали градиентом 120 мл от 0 до 200 мМ имидазола со скоростью 2 мл/мин. Для определения того, какие фракции являются флуоресцентными, использовали коробку с синим светом. Флуоресцентные фракции объединяли и концентрировали до 2–3 мл. Затем белок подвергали диализу с минимум 1 л PBS в течение не менее 24 часов.Среда для автоиндукции состояла из ~900 мл среды ZY (1% триптона и 0,5% дрожжевого экстракта) с 20 мл 50×М (25 мМ Na 2 HPO 4 , 25 мМ KH 2 PO 4 , 50 мМ NH 4 Cl и 5 мМ Na 2 SO 4 ), 20 мл 50 × 5052 (0,5% глицерин, 0,05% глюкоза, 0,2% 0,05% глюкоза, 0,2% 0,8 М α-лактоза), 20 мл 4 , а 200 мкл 1000 × следовые металлы (50 мм FECL 3 , 20 мм CaCl 2 , 10 мм MNCL 2 , 10 мм ZnSO 4 , 2 мм COCL 2 , 2 мм CUCL 2 , 2 мм NICL 2 , 2 мм Na 2 MOO 4 , 2 мм Na 2 SEO 3 , 2 мм H 3 BO 3 ).
Белки очищали с помощью аффинной хроматографии на Ni-NTA-агарозе (колонка 5 мл). Белки iATPSnFR элюировали градиентом 120 мл от 0 до 200 мМ имидазола со скоростью 2 мл/мин. Для определения того, какие фракции являются флуоресцентными, использовали коробку с синим светом. Флуоресцентные фракции объединяли и концентрировали до 2–3 мл. Затем белок подвергали диализу с минимум 1 л PBS в течение не менее 24 часов.Среда для автоиндукции состояла из ~900 мл среды ZY (1% триптона и 0,5% дрожжевого экстракта) с 20 мл 50×М (25 мМ Na 2 HPO 4 , 25 мМ KH 2 PO 4 , 50 мМ NH 4 Cl и 5 мМ Na 2 SO 4 ), 20 мл 50 × 5052 (0,5% глицерин, 0,05% глюкоза, 0,2% 0,05% глюкоза, 0,2% 0,8 М α-лактоза), 20 мл 4 , а 200 мкл 1000 × следовые металлы (50 мм FECL 3 , 20 мм CaCl 2 , 10 мм MNCL 2 , 10 мм ZnSO 4 , 2 мм COCL 2 , 2 мм CUCL 2 , 2 мм NICL 2 , 2 мм Na 2 MOO 4 , 2 мм Na 2 SEO 3 , 2 мм H 3 BO 3 ).
Титрование белков с помощью АТФ
После экспрессии белка 100 мкл белка переносили в черные 96-луночные планшеты с плоским дном (Greiner). Исходную флуоресценцию анализировали на планшетном флуориметре Safire2 со стопером (Tecan). Длины волн возбуждения/испускания составляли 485/515 нм. Усиление составляло от 80 до 120 В, а полосовые фильтры были 5 нм. Флуоресценцию снова анализировали после добавления 10 мкл раствора АТФ, содержащего от 315 нМ до 100 мМ АТФ, для конечных концентраций АТФ, равных 31.5 нМ–10 мМ. После второго чтения было рассчитано d F / F .
Для экспериментов, проведенных с осветленным лизатом HEK-293, чашки диаметром 10 см трансфицировали интересующим цитоплазматическим геном. Через 24 ч после трансфекции клетки дважды промывали PBS. Третью промывку PBS использовали для отделения клеток. Затем клетки осаждали и белок экстрагировали путем лизиса замораживанием/оттаиванием. pH лизата регулировали соответствующим образом, и лизат концентрировали до 1. 5 мл с использованием центробежного фильтра Amicon. После концентрирования лизата строили кривые доза-реакция, как указано выше.
5 мл с использованием центробежного фильтра Amicon. После концентрирования лизата строили кривые доза-реакция, как указано выше.
Для определения рН-зависимости флуоресценции готовили буферные растворы определенного рН (Hydrion Chemvelope) и концентрировали, добавляли очищенный белок до конечной концентрации 0,2 мкМ. Флуоресценцию 100-мкл образцов в трех экземплярах измеряли, как описано выше. Добавляли АТФ (из запаса 200 мМ Mg АТФ) до конечной концентрации 6 мМ и снова измеряли флуоресценцию.
Остановленные кинетики проживания
равные объемы 0,2 мкм очищенный белок в MG -содержащий буфер (10 мМ HEPES, 145 мм NaCl, 2,5 мм KCL, 10 мм глюкоза, 2 мм CaCl 2 , 1 мм MGCL 2 , pH 7,4) и АТФ-содержащий буфер (различные концентрации) смешивали во флуориметре с остановленным потоком SX-20 (Applied Photophysics) с точками данных, собираемыми каждую миллисекунду. Возбуждение осуществлялось от светодиода с длиной волны 490 нм. Эмиссия регистрировалась с помощью фотоумножителя и длинноволнового фильтра 510 нм. Реакции смешения проводили в пяти повторах и усредняли. Данные соответствовали уравнению, содержащему два экспоненциальных роста:
Реакции смешения проводили в пяти повторах и усредняли. Данные соответствовали уравнению, содержащему два экспоненциальных роста:
F=F0+ΔFmax1*1-exp-kobs1*время+ΔFmax2*1-exp-kobs2*время
Мутагенез
Варианты линкера и точечные мутации были получены по сайту -направленный мутагенез по Кункелю 57 с урациловой матрицей. Клетки CJ236 трансформировали многообещающими векторами и выращивали на чашках, содержащих хлорамфеникол, в течение ночи при 37 °C. На следующий день 1 мл культуры LB, содержащей только ампициллин, инокулировали колонией и выращивали при 37 °C при встряхивании до помутнения – обычно около 6 часов.Как только культура становилась мутной, добавляли 1 мкл хелперного фага M13KO7 (NEB: N0315S). После инкубации в течение дополнительного часа культуру размножали до 40 мл культуры LB с ампициллином (без хлорамфеникола) в колбе на 250 мл и инкубировали в течение ночи при 37°С при встряхивании. На следующий день бактерии осаждали при 4000 г в течение 20 мин при 4°С. Супернатант переносили в новую пробирку Falcon объемом 50 мл, содержащую 10 мл 20% ПЭГ в 2,5 М NaCl. После тщательного перемешивания раствор инкубировали на льду в течение 15 мин.Затем фаг осаждали при 4000 г в течение 20 мин. Супернатант удаляли, а фаг ресуспендировали в 2 мл PBS. 2 мл PBS разделяли на две разные пробирки Эппендорфа и центрифугировали на максимальной скорости в течение 5 минут для осаждения оставшихся бактерий. Супернатант переносили в две новые пробирки Эппендорфа, содержащие 300 мкл 20% ПЭГ в 2,5 М NaCl. После встряхивания раствор инкубировали на льду в течение 10 мин. Фаг осаждали на максимальной скорости в течение 5 мин. Супернатант удаляли.Был сделан еще один быстрый отжим для удаления любого остаточного супернатанта. Затем фаг ресуспендировали в 1 мл PBS. Его центрифугировали на максимальной скорости в течение 5 минут, а супернатант переносили в новую пробирку Эппендорфа. Затем урациловую матрицу очищали с использованием набора Qiagen QIAprep Spin M13 Kit (кат.
Супернатант переносили в новую пробирку Falcon объемом 50 мл, содержащую 10 мл 20% ПЭГ в 2,5 М NaCl. После тщательного перемешивания раствор инкубировали на льду в течение 15 мин.Затем фаг осаждали при 4000 г в течение 20 мин. Супернатант удаляли, а фаг ресуспендировали в 2 мл PBS. 2 мл PBS разделяли на две разные пробирки Эппендорфа и центрифугировали на максимальной скорости в течение 5 минут для осаждения оставшихся бактерий. Супернатант переносили в две новые пробирки Эппендорфа, содержащие 300 мкл 20% ПЭГ в 2,5 М NaCl. После встряхивания раствор инкубировали на льду в течение 10 мин. Фаг осаждали на максимальной скорости в течение 5 мин. Супернатант удаляли.Был сделан еще один быстрый отжим для удаления любого остаточного супернатанта. Затем фаг ресуспендировали в 1 мл PBS. Его центрифугировали на максимальной скорости в течение 5 минут, а супернатант переносили в новую пробирку Эппендорфа. Затем урациловую матрицу очищали с использованием набора Qiagen QIAprep Spin M13 Kit (кат. № 27704) по протоколу производителя. 11 мкл буфера MP добавляли к 1 мл раствора PBS/матрицы. После встряхивания смесь инкубировали при комнатной температуре в течение 5 мин. Затем эту смесь наносили на спин-колонку QIAprep и центрифугировали при 4700× g .Проток был отброшен. После нанесения всей смеси на колонку наносили 700 мкл буфера PB. Через 1 мин инкубации колонку центрифугировали при 4700× g в течение 15 с. Это повторяли перед трехкратной промывкой колонки 700 мкл РЕ-буфера и снова центрифугированием при 7000 об/мин в течение 15 с. Убедившись, что колонка сухая, на колонку наносили 25 мкл ЭБ, которую затем центрифугировали при 4700× г . Это было сделано три раза для в общей сложности 75 мкл ЭБ.Качество ДНК проверяли с помощью электрофореза в агарозном геле. Высококачественные препараты представляют собой одну полосу размером около 1/3 размера плазмиды.
№ 27704) по протоколу производителя. 11 мкл буфера MP добавляли к 1 мл раствора PBS/матрицы. После встряхивания смесь инкубировали при комнатной температуре в течение 5 мин. Затем эту смесь наносили на спин-колонку QIAprep и центрифугировали при 4700× g .Проток был отброшен. После нанесения всей смеси на колонку наносили 700 мкл буфера PB. Через 1 мин инкубации колонку центрифугировали при 4700× g в течение 15 с. Это повторяли перед трехкратной промывкой колонки 700 мкл РЕ-буфера и снова центрифугированием при 7000 об/мин в течение 15 с. Убедившись, что колонка сухая, на колонку наносили 25 мкл ЭБ, которую затем центрифугировали при 4700× г . Это было сделано три раза для в общей сложности 75 мкл ЭБ.Качество ДНК проверяли с помощью электрофореза в агарозном геле. Высококачественные препараты представляют собой одну полосу размером около 1/3 размера плазмиды.
Вырожденные праймеры были разработаны для одновременной мутации двух аминокислот. Это было сделано путем фланкирования вырожденной/мутированной последовательности ДНК примерно 15 основаниями немутированной ДНК с каждой стороны. Вариант QUEEN с заменой cpEGFP cpSFGFP был клонирован с помощью ПЦР. Было проведено несколько циклов мутагенеза для скрининга вариантов с мутациями на стыке PS3 и cpSFGFP (линкер 1) и cpSFGFP и PS3 (линкер 2).В следующих праймерах X обозначает кодон, который был рандомизирован для кодирования всех возможных аминокислот. Праймер линкера 2, кодирующий GHKLEYNXXDIDFKRA, привел к варианту с последовательностью линкера 2 GHKLEYNLHDIDFKRA. Это было повторно рандомизировано с рядом праймеров (многие из которых не дали улучшенных сенсоров), включая праймер, кодирующий GHKLEYNFXLHXIDFKRA, из которого была идентифицирована окончательная последовательность линкера 2 GHKLEYNFGLHDIDFKRA. На этом шаблоне линкер 1 был оптимизирован с помощью праймера, кодирующего AERRLQSQXXSHNVYIT.При проверке нескольких сотен вариантов была идентифицирована победная (и окончательная iATPSnFR 1.0 ) последовательность AERRLQSQVLSHNVYIT. Праймеры фосфорилировали с использованием киназы Т4. Реакцию проводили в пробирке Эппендорф, содержащей 18 мкл H 2 O, 3 мкл буфера для киназы Т4, 1 мкл 10 мкМ АТФ, 1 мкл киназы Т4 (NEB M0201L) и 7 мкл 100 мкМ олигоматрицы.
Вариант QUEEN с заменой cpEGFP cpSFGFP был клонирован с помощью ПЦР. Было проведено несколько циклов мутагенеза для скрининга вариантов с мутациями на стыке PS3 и cpSFGFP (линкер 1) и cpSFGFP и PS3 (линкер 2).В следующих праймерах X обозначает кодон, который был рандомизирован для кодирования всех возможных аминокислот. Праймер линкера 2, кодирующий GHKLEYNXXDIDFKRA, привел к варианту с последовательностью линкера 2 GHKLEYNLHDIDFKRA. Это было повторно рандомизировано с рядом праймеров (многие из которых не дали улучшенных сенсоров), включая праймер, кодирующий GHKLEYNFXLHXIDFKRA, из которого была идентифицирована окончательная последовательность линкера 2 GHKLEYNFGLHDIDFKRA. На этом шаблоне линкер 1 был оптимизирован с помощью праймера, кодирующего AERRLQSQXXSHNVYIT.При проверке нескольких сотен вариантов была идентифицирована победная (и окончательная iATPSnFR 1.0 ) последовательность AERRLQSQVLSHNVYIT. Праймеры фосфорилировали с использованием киназы Т4. Реакцию проводили в пробирке Эппендорф, содержащей 18 мкл H 2 O, 3 мкл буфера для киназы Т4, 1 мкл 10 мкМ АТФ, 1 мкл киназы Т4 (NEB M0201L) и 7 мкл 100 мкМ олигоматрицы. Затем реакцию инкубировали в течение 30 мин при 37°С. Чтобы определить оптимальное соотношение матрицы и праймера, первоначально были проведены четыре реакции отжига с использованием четырех различных разведений праймера с киназой; 1:1, 1:5, 1:10 и 1:100.1 мкл разбавленного праймера, 1 мкл матрицы и 1 мкл 10× буфера для ДНК-лигазы Т4 и 7 мкл воды без нуклеазы объединяли в пробирке для ПЦР. Реакционную смесь нагревали до 95°С в течение 2 мин с помощью ПЦР-машины. Затем реакционную смесь медленно доводили до 25°С, используя шаги по 5°С по 30 с каждый. Для полимеризации ДНК: 9,5 мкл воды, 1,5 мкл буфера 10× ДНК-лигазы Т4, 1 мкл 25 мМ dNTP, 1 мкл 10 мМ АТФ, 1 мкл немодифицированной полимеразы Т7 (NEB: M0274L), 1 мкл лигазы Т4 (NEB: M0202M) , и все 10 мкл реакционной смеси для отжига добавляли в пробирку для ПЦР и инкубировали при комнатной температуре в течение часа.Эту реакцию трансформировали непосредственно в клетки E. coli BL21(DE3) (без pLysS) и высевали на чашки LB-Agar размером 24 см × 24 см с 100 мкг/мл ампициллина.
Затем реакцию инкубировали в течение 30 мин при 37°С. Чтобы определить оптимальное соотношение матрицы и праймера, первоначально были проведены четыре реакции отжига с использованием четырех различных разведений праймера с киназой; 1:1, 1:5, 1:10 и 1:100.1 мкл разбавленного праймера, 1 мкл матрицы и 1 мкл 10× буфера для ДНК-лигазы Т4 и 7 мкл воды без нуклеазы объединяли в пробирке для ПЦР. Реакционную смесь нагревали до 95°С в течение 2 мин с помощью ПЦР-машины. Затем реакционную смесь медленно доводили до 25°С, используя шаги по 5°С по 30 с каждый. Для полимеризации ДНК: 9,5 мкл воды, 1,5 мкл буфера 10× ДНК-лигазы Т4, 1 мкл 25 мМ dNTP, 1 мкл 10 мМ АТФ, 1 мкл немодифицированной полимеразы Т7 (NEB: M0274L), 1 мкл лигазы Т4 (NEB: M0202M) , и все 10 мкл реакционной смеси для отжига добавляли в пробирку для ПЦР и инкубировали при комнатной температуре в течение часа.Эту реакцию трансформировали непосредственно в клетки E. coli BL21(DE3) (без pLysS) и высевали на чашки LB-Agar размером 24 см × 24 см с 100 мкг/мл ампициллина. После определения оптимальной концентрации киназных олигонуклеотидов это разведение можно использовать для последующих реакций с этим конкретным урациловым шаблоном.
После определения оптимальной концентрации киназных олигонуклеотидов это разведение можно использовать для последующих реакций с этим конкретным урациловым шаблоном.
Высокопроизводительный скрининг
Отдельные колонии отбирали с помощью робота для сбора колоний QPix2 xt (Genetix). Колонии помещали в глубокие 96-луночные планшеты, заполненные 800 мкл аутоиндукционной среды с 100 мкг/мл ампициллина.Культуры выращивали при энергичном встряхивании в течение ночи при 30 °С. Бактерии собирали центрифугированием (3000 г ) и трижды промывали 900 мкл PBS перед замораживанием. Белок экстрагировали методом замораживания-оттаивания с использованием 900 мкл фосфатно-солевого буфера. Для гомогенизации раствора использовали быстрое встряхивание. Неочищенный лизат осветляли центрифугированием при 4000 g в течение 30 мин. 100 мкл осветленного лизата переносили в плоскодонные черные 96-луночные планшеты (Greiner) с помощью робота для работы с жидкостями. Исходную флуоресценцию анализировали на планшетном флуориметре Safire2 со стопером (Tecan). Длины волн возбуждения/испускания составляли 485/515 нм. Усиление составляло от 80 до 120 В, а полосовые фильтры были 5 нм. Флуоресценцию снова анализировали после добавления 10 мкл 100 мМ АТФ (конечная концентрация АТФ 10 мМ). После расчета d F / F для многообещающих кандидатов были изготовлены новые матрицы урацила, которые были подвергнуты последующим раундам мутагенеза. Наиболее многообещающие сенсоры были секвенированы и клонированы либо в pDisplay, либо в pDisplayMini для дальнейшего скрининга в клетках млекопитающих.
Исходную флуоресценцию анализировали на планшетном флуориметре Safire2 со стопером (Tecan). Длины волн возбуждения/испускания составляли 485/515 нм. Усиление составляло от 80 до 120 В, а полосовые фильтры были 5 нм. Флуоресценцию снова анализировали после добавления 10 мкл 100 мМ АТФ (конечная концентрация АТФ 10 мМ). После расчета d F / F для многообещающих кандидатов были изготовлены новые матрицы урацила, которые были подвергнуты последующим раундам мутагенеза. Наиболее многообещающие сенсоры были секвенированы и клонированы либо в pDisplay, либо в pDisplayMini для дальнейшего скрининга в клетках млекопитающих.
Трансфекция клеток HEK293 и U373
Клетки HEK293 (ATCC, CRL-1573) и U373MG (Sigma-Aldrich, 08061901) поддерживали в среде DMEM/F-12 с добавлением GlutaMAX (Gibco: 10565042), 10% FBS ( Gibco 10082147) и 5 мл пенициллина-стрептомицина (10 000 ЕД/мл) (Gibco: 15140-122) при 37 °C с 5% CO 2 . Клетки обрабатывали трипсином, как только они достигали слияния, и разводили в соотношении 1:10. Клетки HEK-293 и U373MG трансфицировали либо липофектамином 2000 (ThermoFisher 11668027), либо эффектеном (Qiagen ID: 301425) в соответствии с рекомендациями производителя.Ниже приведена процедура трансфекции одной лунки 6-луночной чашки. 100 мкл OptiMEM (Gibco: 31985062) помещали в две пробирки. В одну пробирку добавляли 2,5 мкг ДНК, а в другую — 6,25 мкл липофектамина 2000. Содержимое двух пробирок объединяли, тщательно перемешивали и инкубировали в течение 20 мин при комнатной температуре перед добавлением смеси к клеткам, конфлюэнтным на 70–90% со свежей средой. Среду меняли через 6–8 ч. В пробирку добавляли 0,4 мкг ДНК, а затем 8 мкл энхансера.100 мкл пипетированного буфера ЕС использовали для смешивания содержимого пипетированием. Затем смесь инкубировали в течение 2 минут, прежде чем тщательно смешать с ней 8 мкл Effectene. Затем раствор инкубировали в течение 10 мин. 600 мкл свежей среды добавляли перед добавлением смеси к клеткам, которые были на 70–90% конфлюэнтны со свежей средой.
Клетки HEK-293 и U373MG трансфицировали либо липофектамином 2000 (ThermoFisher 11668027), либо эффектеном (Qiagen ID: 301425) в соответствии с рекомендациями производителя.Ниже приведена процедура трансфекции одной лунки 6-луночной чашки. 100 мкл OptiMEM (Gibco: 31985062) помещали в две пробирки. В одну пробирку добавляли 2,5 мкг ДНК, а в другую — 6,25 мкл липофектамина 2000. Содержимое двух пробирок объединяли, тщательно перемешивали и инкубировали в течение 20 мин при комнатной температуре перед добавлением смеси к клеткам, конфлюэнтным на 70–90% со свежей средой. Среду меняли через 6–8 ч. В пробирку добавляли 0,4 мкг ДНК, а затем 8 мкл энхансера.100 мкл пипетированного буфера ЕС использовали для смешивания содержимого пипетированием. Затем смесь инкубировали в течение 2 минут, прежде чем тщательно смешать с ней 8 мкл Effectene. Затем раствор инкубировали в течение 10 мин. 600 мкл свежей среды добавляли перед добавлением смеси к клеткам, которые были на 70–90% конфлюэнтны со свежей средой. СМИ были заменены на следующий день.
СМИ были заменены на следующий день.
Лизис клеток НЕК293
Клетки НЕК293 трижды промывали холодным PBS перед лизисом клеток. Клетки лизировали на льду с использованием буфера RIPA: 150 мМ NaCl, 20 мМ Трис, 1 мМ ЭДТА, 1 % Тритон, 0.1% SDS и 0,5% дезоксихолата натрия при pH 8 с добавлением ингибиторов протеазы (ThermoFisher). После 10-минутной инкубации на льду смесь центрифугировали при максимальной скорости в течение 10-минут. Супернатант брали и разбавляли равным объемом 2× буфера Лэммли для вестерн-блоттинга.
Вестерн-блоттинг
Готовые гели были приобретены в компании BioRad и подвергались анализу в течение 110 мин при 100 В. 1× буфер для прогонки состоял из 3,0 г трис-основания, 14,4 г глицина и 1,0 г додецилсульфата натрия в 1 л воды.Гели переносили на нитроцеллюлозу на 1 ч при 100 В в буфере для переноса 1×. 10-кратный буфер для переноса состоял из 30,0 г Триса и 144,0 г глицина в 1 л воды. 1× буфер для переноса состоит из 10% рабочего буфера 10×, 20% метанола и 70% воды. После переноса блоты блокировали 5% бычьим сывороточным альбумином (BSA) (Sigma: A9647-100G) в TBS с 0,05% Tween-20 (TBST) (Sigma: P2287-100ML) в течение 1 часа. Антитела получали с 5% BSA в TBST. Первичные антитела: мышь GAPDH 1:5000 (Thermofisher MA5-15738).GFP 1:1000 кролик (Invitrogen A11122). Вторичные антитела: IRDye 680RD антимышиные 1:20000 (Licor 926-68170). IRDye 800CW против кролика 1:20 000 (Licor 925-68024). Первичные антитела инкубировали в течение ночи при 4°С. После трех промывок TBST продолжительностью не менее 10 мин каждая вторичные антитела инкубировали в течение 1 ч при комнатной температуре. Трех промывок TBST продолжительностью не менее 10 мин каждая было достаточно для удаления вторичных антител. Мембраны визуализировали с помощью тепловизора Licor Odyssey.
После переноса блоты блокировали 5% бычьим сывороточным альбумином (BSA) (Sigma: A9647-100G) в TBS с 0,05% Tween-20 (TBST) (Sigma: P2287-100ML) в течение 1 часа. Антитела получали с 5% BSA в TBST. Первичные антитела: мышь GAPDH 1:5000 (Thermofisher MA5-15738).GFP 1:1000 кролик (Invitrogen A11122). Вторичные антитела: IRDye 680RD антимышиные 1:20000 (Licor 926-68170). IRDye 800CW против кролика 1:20 000 (Licor 925-68024). Первичные антитела инкубировали в течение ночи при 4°С. После трех промывок TBST продолжительностью не менее 10 мин каждая вторичные антитела инкубировали в течение 1 ч при комнатной температуре. Трех промывок TBST продолжительностью не менее 10 мин каждая было достаточно для удаления вторичных антител. Мембраны визуализировали с помощью тепловизора Licor Odyssey.
Микроинъекции AAV2/5 in vivo
Все эксперименты на животных проводились в соответствии с Руководством Национального института здравоохранения по уходу и использованию лабораторных животных и были одобрены Комитетом по исследованиям на животных канцлера Калифорнийского университета в Лос-Анджелесе. Мыши C57BL/6NTac дикого типа были получены из колоний домашнего разведения или приобретены у Taconic Biosciences. Всех мышей содержали с пищей и водой без ограничений в 12-часовой светлой/темной среде. Все животные были здоровы, без явного поведенческого фенотипа, не участвовали в предыдущих исследованиях и были умерщвлены во время светового цикла. Первоначально животных глубоко анестезировали в камере с 5% изофлураном. Затем голову животного брили, чтобы удалить мех вокруг хирургического участка.Голову животного фиксировали в стереотаксической установке с помощью ушных перекладин. На нос помещали носовой конус для непрерывного введения изофлурана на протяжении всей процедуры. Проводился постоянный мониторинг дыхания животного во время процедуры и соответствующим образом корректировалась анестезия (0,5–3%). Животному вводили бупренорфин (0,1 мг/кг бупренекса), а перед операцией место хирургического вмешательства трижды очищали 10% повидон-йодом и 70% этанолом. Был сделан небольшой разрез, чтобы обнажить череп.
Мыши C57BL/6NTac дикого типа были получены из колоний домашнего разведения или приобретены у Taconic Biosciences. Всех мышей содержали с пищей и водой без ограничений в 12-часовой светлой/темной среде. Все животные были здоровы, без явного поведенческого фенотипа, не участвовали в предыдущих исследованиях и были умерщвлены во время светового цикла. Первоначально животных глубоко анестезировали в камере с 5% изофлураном. Затем голову животного брили, чтобы удалить мех вокруг хирургического участка.Голову животного фиксировали в стереотаксической установке с помощью ушных перекладин. На нос помещали носовой конус для непрерывного введения изофлурана на протяжении всей процедуры. Проводился постоянный мониторинг дыхания животного во время процедуры и соответствующим образом корректировалась анестезия (0,5–3%). Животному вводили бупренорфин (0,1 мг/кг бупренекса), а перед операцией место хирургического вмешательства трижды очищали 10% повидон-йодом и 70% этанолом. Был сделан небольшой разрез, чтобы обнажить череп. С помощью высокоскоростной дрели в черепе было просверлено отверстие ~2 мм на расстоянии 2 мм кзади от брегмы и 1,5 мм слева от брегмы для гиппокампа СА1, а также на расстоянии 0,8 мм кпереди от брегмы и 2 мм слева от брегмы и для дорсолатерального полосатого тела. После удаления части черепа на место хирургического вмешательства наносили физиологический раствор, чтобы удалить любой мусор. В область СА1 гиппокампа вводили 1 мкл вируса (титр 1,5Е13) в течение 5 мин. Используемые стереотаксические координаты: 2 мм кзади от брегмы, 1,5 мм левее средней линии и 1.6 мм от пиальной поверхности для гиппокампа СА1 и 0,8 мм кпереди от брегмы, 2 мм слева от средней линии и 2,4 мм от пиальной поверхности для дорсолатерального полосатого тела. После завершения инъекции игла оставалась на месте в течение 10 минут, чтобы позволить вирусу диффундировать в ткани. Игла постепенно удалялась в течение 2 мин. Операционное поле было ушито. Животные выздоровели в клетке, помещенной на грелку. На следующий день животным давали 0,1 мг/кг бупренекса от боли.
С помощью высокоскоростной дрели в черепе было просверлено отверстие ~2 мм на расстоянии 2 мм кзади от брегмы и 1,5 мм слева от брегмы для гиппокампа СА1, а также на расстоянии 0,8 мм кпереди от брегмы и 2 мм слева от брегмы и для дорсолатерального полосатого тела. После удаления части черепа на место хирургического вмешательства наносили физиологический раствор, чтобы удалить любой мусор. В область СА1 гиппокампа вводили 1 мкл вируса (титр 1,5Е13) в течение 5 мин. Используемые стереотаксические координаты: 2 мм кзади от брегмы, 1,5 мм левее средней линии и 1.6 мм от пиальной поверхности для гиппокампа СА1 и 0,8 мм кпереди от брегмы, 2 мм слева от средней линии и 2,4 мм от пиальной поверхности для дорсолатерального полосатого тела. После завершения инъекции игла оставалась на месте в течение 10 минут, чтобы позволить вирусу диффундировать в ткани. Игла постепенно удалялась в течение 2 мин. Операционное поле было ушито. Животные выздоровели в клетке, помещенной на грелку. На следующий день животным давали 0,1 мг/кг бупренекса от боли. После выздоровления за животными наблюдали, чтобы убедиться, что они здоровы, и они получали триметоприм сульфаметоксазол с пищей в течение недели после операции.Нацеливание на MSN было достигнуто с использованием FLEX-зависимого iATPSnFR 1,0 AAV с селективными линиями мышей Cre 58 D1 и D2.
После выздоровления за животными наблюдали, чтобы убедиться, что они здоровы, и они получали триметоприм сульфаметоксазол с пищей в течение недели после операции.Нацеливание на MSN было достигнуто с использованием FLEX-зависимого iATPSnFR 1,0 AAV с селективными линиями мышей Cre 58 D1 и D2.
Визуализация
Клетки HEK293 временно трансфицировали вектором, кодирующим лидерную последовательность IgK, за которой следовали iATPSnFR, линкер GGTGGS, метка myc и трансмембранный домен рецептора PDGF. После 24 ч инкубации при 37°С с 5% СО 2 в 6-луночных планшетах клетки обрабатывали трипсином и высевали на покровные стекла, предварительно покрытые полилизином.После еще 24-часовой инкубации при 37°C с 5% CO 2 покровные стекла помещали в перфузионную камеру. Изображения были получены с помощью Olympus IXS 71 с использованием масляной иммерсионной линзы UplansApo 40 × 0,90 NA или конфокального микроскопа Olympus BX51WI с иммерсионной линзой LUMPlanFL/IR 40 × 0,80 NA. После помещения в перфузионную камеру клетки перфузировали постоянным потоком свежего буфера HEK293 до и после нанесения лигандов. Растворы АТФ готовили ежедневно. Стационарный уровень флуоресценции рассчитывали как среднее значение изображений, полученных за первые 30 с.Быстрые изменения решения были достигнуты с помощью быстродействующего пошагового контроллера перфузии SF-77B от Warner Instruments. Стеклянный квадрат с тремя цилиндрами (Warner cat 64-0119) помещали как можно ближе к ячейке для целей визуализации. Шаги 800 мкм использовались для перемещения бочек на клетки и от них. Скорость включения и выключения определяли с помощью pClamp 10.6. Буфер HEK: 140 мМ NaCl, 1 мМ MgCl 2 , 2 мМ CaCl 2 , 10 мМ HEPES и 10 мМ глюкозы, доведенной до pH 7,4 с помощью 5 мМ NaOH. Оценку внутриклеточного pH проводили с использованием BCECF-AM в соответствии с инструкциями производителя (Invitrogen).Для конфокальной микроскопии мы использовали возбуждение 473 нм и эмиссию 525 нм.
После помещения в перфузионную камеру клетки перфузировали постоянным потоком свежего буфера HEK293 до и после нанесения лигандов. Растворы АТФ готовили ежедневно. Стационарный уровень флуоресценции рассчитывали как среднее значение изображений, полученных за первые 30 с.Быстрые изменения решения были достигнуты с помощью быстродействующего пошагового контроллера перфузии SF-77B от Warner Instruments. Стеклянный квадрат с тремя цилиндрами (Warner cat 64-0119) помещали как можно ближе к ячейке для целей визуализации. Шаги 800 мкм использовались для перемещения бочек на клетки и от них. Скорость включения и выключения определяли с помощью pClamp 10.6. Буфер HEK: 140 мМ NaCl, 1 мМ MgCl 2 , 2 мМ CaCl 2 , 10 мМ HEPES и 10 мМ глюкозы, доведенной до pH 7,4 с помощью 5 мМ NaOH. Оценку внутриклеточного pH проводили с использованием BCECF-AM в соответствии с инструкциями производителя (Invitrogen).Для конфокальной микроскопии мы использовали возбуждение 473 нм и эмиссию 525 нм. Для эпифлуоресцентной микроскопии мы использовали логометрическую визуализацию с чередующимся возбуждением 440 (500 мс), 505 нм (50 мс) и эмиссионной визуализацией 535 нм. В этих экспериментах использовались монохроматор (Till Photonics Polychrome IV или V) и ПЗС (Till Imago от Till Photonics), при этом управление изображением осуществлялось программным обеспечением TillVision.
Для эпифлуоресцентной микроскопии мы использовали логометрическую визуализацию с чередующимся возбуждением 440 (500 мс), 505 нм (50 мс) и эмиссионной визуализацией 535 нм. В этих экспериментах использовались монохроматор (Till Photonics Polychrome IV или V) и ПЗС (Till Imago от Till Photonics), при этом управление изображением осуществлялось программным обеспечением TillVision.
Животных глубоко анестезировали пентобарбиталом (внутрибрюшинно) перед транскардиальной перфузией.При отсутствии рефлексов вскрывали грудную полость. Левый желудочек прокалывали иглой, соединенной с перфузионным насосом, а правый желудочек пунктировали для обеспечения транскардиальной перфузии. Животным вводили не менее 30 мл охлажденного льдом PBS для полного смыва крови, а затем вводили не менее 30 мл 10% забуференного формалина для фиксации тканей. Ткани аккуратно вырезали и фиксировали в течение ночи в 10% забуференном формалине при 4 °C. Затем ткани подвергали криозащите с помощью 30% сахарозы в PBS, а затем замораживали в O. КТ соединения (Fisher Scientific: 23-730-571). Срезы толщиной 40 мкм собирали с помощью криостатного микротома (Leica). Срезы собирали в PBS и блокировали 10% нормальной козьей сывороткой и 0,5% Triton-X 100 в PBS на 1 ч при комнатной температуре перед окрашиванием. Первичное антитело наносили на ночь при 4°C в PBS с 10% нормальной козьей сывороткой и 0,5% Triton-X 100. После первичного нанесения срезы трижды промывали PBS. Вторичное антитело инкубировали при комнатной температуре в течение 1 часа с последующим трехкратным промыванием PBS перед нанесением на предметные стекла.Срезы полностью высохли на предметном стекле перед нанесением Fluoromount-G (SouthernBiotech: 0100-01). Первичные антитела: S100β кролика 1:1000 (Abcam ab41548). Кролик NeuN 1:1000 (Cell Signaling D3S3I). Цыпленок GFP 1:1000 (Abcam ab13970). Вторичные антитела: козий антикуриный IgG Alexa 488 1:2000 (Invitrogen A11039). Коза против кролика Alexa 546 1:2000 (Invitrogen A11010). Для визуализации красителей Alexa 488 использовался аргоновый лазер с длиной волны 488 нм, отрегулированный на максимальную выходную интенсивность 5–10%.
КТ соединения (Fisher Scientific: 23-730-571). Срезы толщиной 40 мкм собирали с помощью криостатного микротома (Leica). Срезы собирали в PBS и блокировали 10% нормальной козьей сывороткой и 0,5% Triton-X 100 в PBS на 1 ч при комнатной температуре перед окрашиванием. Первичное антитело наносили на ночь при 4°C в PBS с 10% нормальной козьей сывороткой и 0,5% Triton-X 100. После первичного нанесения срезы трижды промывали PBS. Вторичное антитело инкубировали при комнатной температуре в течение 1 часа с последующим трехкратным промыванием PBS перед нанесением на предметные стекла.Срезы полностью высохли на предметном стекле перед нанесением Fluoromount-G (SouthernBiotech: 0100-01). Первичные антитела: S100β кролика 1:1000 (Abcam ab41548). Кролик NeuN 1:1000 (Cell Signaling D3S3I). Цыпленок GFP 1:1000 (Abcam ab13970). Вторичные антитела: козий антикуриный IgG Alexa 488 1:2000 (Invitrogen A11039). Коза против кролика Alexa 546 1:2000 (Invitrogen A11010). Для визуализации красителей Alexa 488 использовался аргоновый лазер с длиной волны 488 нм, отрегулированный на максимальную выходную интенсивность 5–10%. HeNe-лазер 543, настроенный на максимальную выходную интенсивность 20–30%, использовался для визуализации красителей Alexa 546.
HeNe-лазер 543, настроенный на максимальную выходную интенсивность 20–30%, использовался для визуализации красителей Alexa 546.
Съемник пипеток модели P-97 от Sutter Instrument Co. использовался для извлечения капилляров, приобретенных у VWR (1B100-4). Пипетки с пластырями заполняли 3 мМ АТФ и гидразидом Alexa Fluor 568 (1:200) (ThermoFisher Scientific: A10441). С помощью микроманипуляторов кончик пипетки помещали на 10–20 мкм выше изображаемой клетки. Постоянный поток свежего буфера все время заполнял камеру визуализации. После 30-секундной визуализации через PicoSpritzer III от Intracel вводили 5-секундную затяжку 3 мМ АТФ.
Перед обезглавливанием животных глубоко анестезировали изофлураном. Мозг извлекали как можно бережнее и быстрее, а затем как можно быстрее охлаждали. Мозг прикрепляли суперклеем к режущей платформе и заливали ледяным буфером для срезов гиппокампа, в то время как срезы толщиной 300 мкм собирали с помощью вибратома. Для срезов спинного мозга выполняли ламинэктомию. Твердая мозговая оболочка и все корешки были удалены, после чего спинной мозг с помощью суперклея был смонтирован на агарозном блоке для поперечного среза (300 мкм).Затем эти срезы помещали в буфер для визуализации гиппокампа при температуре 32°C. Все растворы барботировали 95% O 2 и 5% CO 2 в течение не менее 30 минут перед нарезкой. После разрезания срезы помещали в перфузионную камеру для визуализации с постоянным потоком 2–3 мл/мин буфера для визуализации гиппокампа. Изображения были получены с использованием конфокального микроскопа Olympus BX51WI с водно-иммерсионным объективом LUMPlanFL/IR 40 × 0,80 NA. Аргоновый лазер с длиной волны 488 нм, настроенный на максимальную выходную интенсивность 10–15% для визуализации GFP.Гиппокампал нарезки буфера: 87 мм NaCl, 25 мм NaHCO 3 , 25 мм глюкоза, 2,5 мм ККК, 1,25 мм Nah 2 PO 4 , 75 мм сахароза, 7 мм MGCL 2 и 0,5 мм CaCl 2 . Гиппокампальный визуализационный буфер: 126 мм NaCl, 25 мМ NaHCO 3 , 10 мМ глюкозы, 2,5 мМ ККЦ, 1,24 мм Nah 2 PO 4 , 1,30 мм MGCL 2 и 2,4 мм CaCl 2 .
Твердая мозговая оболочка и все корешки были удалены, после чего спинной мозг с помощью суперклея был смонтирован на агарозном блоке для поперечного среза (300 мкм).Затем эти срезы помещали в буфер для визуализации гиппокампа при температуре 32°C. Все растворы барботировали 95% O 2 и 5% CO 2 в течение не менее 30 минут перед нарезкой. После разрезания срезы помещали в перфузионную камеру для визуализации с постоянным потоком 2–3 мл/мин буфера для визуализации гиппокампа. Изображения были получены с использованием конфокального микроскопа Olympus BX51WI с водно-иммерсионным объективом LUMPlanFL/IR 40 × 0,80 NA. Аргоновый лазер с длиной волны 488 нм, настроенный на максимальную выходную интенсивность 10–15% для визуализации GFP.Гиппокампал нарезки буфера: 87 мм NaCl, 25 мм NaHCO 3 , 25 мм глюкоза, 2,5 мм ККК, 1,25 мм Nah 2 PO 4 , 75 мм сахароза, 7 мм MGCL 2 и 0,5 мм CaCl 2 . Гиппокампальный визуализационный буфер: 126 мм NaCl, 25 мМ NaHCO 3 , 10 мМ глюкозы, 2,5 мМ ККЦ, 1,24 мм Nah 2 PO 4 , 1,30 мм MGCL 2 и 2,4 мм CaCl 2 .
Анализ данных и статистика
Данные каждого эксперимента представляют минимум n мышей (для экспериментов, в которых использовались мыши).Во всех экспериментах по визуализации n было >3 независимых оценок (мыши или трансфекции). Размеры выборки не рассчитывались априори. На панелях рисунка и в подписях и определяются как количество повторов (клеток, дендритов, интересующих областей или оценок в случае биохимических оценок) и обычно относятся к количеству клеток, если не указано иное. Для инъекций AAV мышей случайным образом распределяли в каждую экспериментальную группу. Никакие точки экспериментальных данных не были исключены.Статистические тесты проводились в Origin 9. Сводные данные представлены как среднее ± sem. Обратите внимание, что на некоторых графиках столбцы, представляющие sem. меньше, чем символы, используемые для представления среднего значения. Для каждого набора сравниваемых данных мы определяли в Origin или GraphPad InStat, были ли данные распределены нормально или нет, используя тест Шапиро-Уилка. Если они были нормально распределены, мы использовали параметрические тесты. Если данные не были нормально распределены, мы использовали непараметрические тесты.Парные и непарные двусторонние критерии t Стьюдента (соответственно) и двусторонние критерии Манна-Уитни использовались для большинства статистических анализов со значимостью, заявленной на уровне p < 0,05. Когда применялся статистический тест, не являющийся критерием t Стьюдента или критерием Манна-Уитни для конкретного случая, то это указывается в тексте как таковое. На протяжении всей рукописи результаты статистических тестов ( p значений и n чисел) полностью представлены на панелях рисунков, чтобы сэкономить место в основной части рукописи.Используются точные значения p , но когда значение p было меньше 0,0001, оно указывается как p < 0,0001, поскольку используемая нами программа не дает точных чисел для таких низких значений. Номера n указаны на панелях рисунков и представляют собой дискретные образцы.
Если они были нормально распределены, мы использовали параметрические тесты. Если данные не были нормально распределены, мы использовали непараметрические тесты.Парные и непарные двусторонние критерии t Стьюдента (соответственно) и двусторонние критерии Манна-Уитни использовались для большинства статистических анализов со значимостью, заявленной на уровне p < 0,05. Когда применялся статистический тест, не являющийся критерием t Стьюдента или критерием Манна-Уитни для конкретного случая, то это указывается в тексте как таковое. На протяжении всей рукописи результаты статистических тестов ( p значений и n чисел) полностью представлены на панелях рисунков, чтобы сэкономить место в основной части рукописи.Используются точные значения p , но когда значение p было меньше 0,0001, оно указывается как p < 0,0001, поскольку используемая нами программа не дает точных чисел для таких низких значений. Номера n указаны на панелях рисунков и представляют собой дискретные образцы. Однако там, где это уместно, в тексте также приводятся основные статистические данные. Изображения были проанализированы с помощью ImageJ. Плагин GECIquant 59 использовался для вычитания фона из изображений, а также для выбора областей интереса.Измерения среднего значения серого были импортированы в Excel для анализа d F / F . Окончательные результаты были представлены в графическом виде в OriginPro 2016, а рисунки собраны в CorelDraw 2017 для создания файлов рисунков в формате TIFF.
Однако там, где это уместно, в тексте также приводятся основные статистические данные. Изображения были проанализированы с помощью ImageJ. Плагин GECIquant 59 использовался для вычитания фона из изображений, а также для выбора областей интереса.Измерения среднего значения серого были импортированы в Excel для анализа d F / F . Окончательные результаты были представлены в графическом виде в OriginPro 2016, а рисунки собраны в CorelDraw 2017 для создания файлов рисунков в формате TIFF.
КАК СМОТРЕТЬ ПРЯМУЮ ТРАНСЛЯЦИЮ NFR РОДЕО ОНЛАЙН
Привет и добро пожаловать
Национальный финал Wrangler Rodeo начинается в четверг, 2 декабря, и Cowboy Channel+ будет там для всех. Турнир из 10 туров продлится с декабря.2-11 в Лас-Вегасе, и в нем примут участие 15 лучших участников во всем, от езды на быках до борьбы с быками. Конкурсанты ищут призовой фонд в размере 10 миллионов долларов в Центре Томаса и Мака в Лас-Вегасе.
| Купон | Купон (скидка | ||
| ковбойский канал (в режиме реального времени) | NFR | ||
| RFD-TV 81180 | NFR | ||
| Cowboy Channel Plus (International) | Часы | NFR | NFR |
Национальные финальные финалы 2020 Rodeo Rodeo (NFR) в прямом эфире в четверг, 12 декабря.2-11 из Центра Томаса и Мака, Лас-Вегас, начало в 22:00. Восточное время/19:00 PT на канале Cowboy +. В главном в мире родео участвуют 14 лучших участников в езде без седла, борьбе с быками, командной езде на канатах, верховой езде в седле, езде на канатах, гонках на бочках и езде на быках. Таким образом, вы не должны пропустить ни одного момента действия NFR.
Национальный финал Родео в прямом эфире на канале Cowboy+
Смотрите трансляции WNFR в прямом эфире каждую ночь на The Cowboy Channel+ с 10:00. м. – 1:00 (ET) каждую ночь дома. Все, что вам нужно, это настроиться на Dish: Channel 158; Прямое телевидение: канал 221. Или проверьте свои местные списки. Вышеуказанные каналы доступны для США, Канады, Мексики, Бразилии, Великобритании, Австралии и всех международных зрителей. Если вы любите смотреть основные моменты родео NFR, то roartube — лучшее место рядом с YouTube, чтобы смотреть национальные финалы родео.
м. – 1:00 (ET) каждую ночь дома. Все, что вам нужно, это настроиться на Dish: Channel 158; Прямое телевидение: канал 221. Или проверьте свои местные списки. Вышеуказанные каналы доступны для США, Канады, Мексики, Бразилии, Великобритании, Австралии и всех международных зрителей. Если вы любите смотреть основные моменты родео NFR, то roartube — лучшее место рядом с YouTube, чтобы смотреть национальные финалы родео.
Раунд 1 национального финала Wrangler
Дата: четверг, 2 декабря | Время: 10 ч.м. ET
Местонахождение: Thomas & Mack Center, Лас-Вегас
Телевидение: Прямая трансляция на канале Cowboy Channel+ в 22:00. ET (ежедневный повторный эфир в 1:00 по восточноевропейскому времени)
Зарегистрироваться на раунд 1
Раунд 2 национального финала Wrangler
Дата: пятница, 3 декабря | Время: 22:00 ET
Местонахождение: Thomas & Mack Center, Лас-Вегас
Телевидение: Прямая трансляция на канале Cowboy Channel+ в 22:00. ET (ежедневный повторный эфир в 1 час.м. ET)
ET (ежедневный повторный эфир в 1 час.м. ET)
ЗАПИСАТЬСЯ на 2-й раунд
3-й раунд национального финала Wrangler
Дата: суббота, 4 декабря | Время: 22:00 ET
Местонахождение: Thomas & Mack Center, Лас-Вегас
Телевидение: Прямая трансляция на канале Cowboy Channel+ в 22:00. ET (ежедневный повтор в 1:00 по восточноевропейскому времени)
Запись на 3-й раунд
4-й раунд национального финала Wrangler
Дата: воскресенье, 5 декабря | Время: 10 ч.м. ET
Местонахождение: Thomas & Mack Center, Лас-Вегас
Телевидение: Прямая трансляция на канале Cowboy Channel+ в 22:00. ET (ежедневный повторный эфир в 1:00 по восточноевропейскому времени)
Запись на 4-й раунд
5-й раунд национального финала Wrangler
Дата: понедельник, 6 декабря | Время: 22:00 ET
Местонахождение: Thomas & Mack Center, Лас-Вегас
Телевидение: Прямая трансляция на канале Cowboy Channel+ в 22:00. ET (ежедневный повторный эфир в 1 час.м. ET )
ET (ежедневный повторный эфир в 1 час.м. ET )
ЗАПИСАТЬСЯ на 5-й раунд
6-й раунд национального финала Wrangler
Дата: вторник, 7 декабря | Время: 22:00 ET
Местонахождение: Thomas & Mack Center, Лас-Вегас
Телевидение: Прямая трансляция на канале Cowboy Channel+ в 22:00. ET (ежедневный повтор в 1:00 по восточноевропейскому времени)
Запись на 6-й раунд
7-й раунд национального финала Wrangler
Дата: среда, 8 декабря | Время: 10 ч.м. ET
Местонахождение: Thomas & Mack Center, Лас-Вегас
Телевидение: Прямая трансляция на канале Cowboy Channel+ в 22:00. ET (ежедневный повтор в 1:00 по восточноевропейскому времени)
Запись на 7-й раунд
8-й раунд национального финала Wrangler
Дата: четверг, 9 декабря | Время: 22:00 ET
Местонахождение: Thomas & Mack Center, Лас-Вегас
Телевидение: Прямая трансляция на канале Cowboy Channel+ в 22:00. ET (ежедневный повторный эфир в 1 час.м. ET)
ET (ежедневный повторный эфир в 1 час.м. ET)
ЗАПИСАТЬСЯ на 8-й раунд
9-й раунд национального финала Wrangler
Дата: пятница, 10 декабря | Время: 22:00 ET
Местонахождение: Thomas & Mack Center, Лас-Вегас
Телевидение: Прямая трансляция на канале Cowboy Channel+ в 22:00. ET (ежедневный повторный эфир в 1:00 по восточноевропейскому времени)
Запись на 9-й раунд
10-й раунд национального финала Wrangler
Дата: суббота, 11 декабря | Время: 10 ч.м. ET
Местонахождение: Thomas & Mack Center, Лас-Вегас
Телевидение: Прямая трансляция на канале Cowboy Channel+ в 22:00. ET (ежедневный повторный эфир в 1:00 по восточноевропейскому времени)
Зарегистрируйтесь для участия в 10-м раунде
Присоединяйтесь к «Ковбойскому Рождеству» в Техасском конференц-центре, South Halls на национальном финале родео, начиная с четверга, 2 декабря, и заканчивая 11 декабря! И не забудьте надеть розовое платье на мероприятие Tough Enough to Wear Pink Night, спонсируемое Montana Silversmiths, которое состоится 11 декабря в Центре Томаса и Мака . Пока вы находитесь в Wrangler NFR, зайдите и сделайте покупки в South Point Western Gift Show в выставочном зале South Point и в зале Boot Barn и обязательно посетите обед Miss Rodeo America PRCA в MGM Grand Conference Center # Room 319- 320.
Пока вы находитесь в Wrangler NFR, зайдите и сделайте покупки в South Point Western Gift Show в выставочном зале South Point и в зале Boot Barn и обязательно посетите обед Miss Rodeo America PRCA в MGM Grand Conference Center # Room 319- 320.
Wrangler NFR проводится каждый декабрь в Центре Томаса и Мака в Лас-Вегасе по контракту с Ассоциацией профессиональных родео-ковбоев (PRCA), который действует до 2024 года, кошелек в 2018 году составляет 10 миллионов долларов. Квалификация основана на мировом зачете PRCA 2018 года.
О Wrangler NFR
Стандартные родео-состязания включают езду без седла, борьбу с быками, командную езду на канатах (хедлеры и хилеры), езду в седлах на бронях, езду на веревках, гонки на бочках WPRA и езду на быках. Это захватывающий вид спорта, который требует терпения, храбрости и просто щепотки безумия, чтобы показать себя хорошо. Чемпион определяется по доходам за сезон. Каждый участник будет выступать один раз в день. В конце каждого раунда в каждом событии есть два чемпиона. Лучшее совокупное время / результат в конечном итоге выиграет событие.
Лучшее совокупное время / результат в конечном итоге выиграет событие.
Cowboy Channel+ предоставляет возможность смотреть Wrangler NFR и другие чемпионаты в течение всего года. Всегда что-то происходит, и это событие, в частности, обещает быть захватывающим по множеству причин. В родео скрыта скрытая опасность, которая делает их интригующими, и следующие 10 дней будут очень интересными для этого события.
Ларсен гордится ранговыми поездками
Орин Ларсен едет на Mr. Harry Стейса Смита Pro Rodeo и занимает место в восьмом заезде в четверг на национальном финале Wrangler Rodeo.
(ФОТО РОББИ ФРИМЭНА)
ЛАС-ВЕГАС – За 12 лет верховой езды на неоседланных лошадях в PRCA Орин Ларсен известен тем, что ездит на высококлассных бронзах.
Он доказал это в четверг, набрав 84,5 балла на Mr. Harry Стейса Смита Pro Rodeo и финишировав четвертым в восьмом круге. Это стоило 11 321 доллар и доказывало, что, когда канадский ковбой набирает обороты, он просто позволяет этому катиться. Он занимает третью ночь подряд и заработал 52 673 доллара, большая часть которых пришлась на 6-8 раунды.
Он занимает третью ночь подряд и заработал 52 673 доллара, большая часть которых пришлась на 6-8 раунды.
«Возможно, я не выигрываю («Eliminator Pen») каждый год, но, похоже, я занимаю довольно хорошие места», — сказал Ларсен из Инглиса, Манитоба, который сейчас живет в Геринге, штат Небраска. «Я просто очень люблю этих действительно сильных лошадей, которые проверят вас и заставят заработать. Когда у вас 90 баллов, это похоже на поездку на 90 баллов.
«Каждый раз, когда вы можете поместить в E-pen в финале, это очень хорошее чувство».
Ларсен занимает пятое место в общем зачете с 677 баллами за восемь поездок.5 баллов. Если он останется на этой позиции, когда NFR завершится в субботу, он добавит бонус в размере 23 513 долларов. Если он переместится на одну позицию вверх, на четвертую, бонус увеличится почти до 33 000 долларов. Он мог использовать любые деньги, которые он мог получить. Он занимает 11-е место в мировом зачете с $150 516.
«Я столкнулся с мистером Гарри в этом раунде два года назад, и я чувствовал то же самое», — сказал он. «Это просто бой. С того момента, как ты киваешь головой, это просто постоянная битва все время. Ричи (Чемпион) связался с ним ранее на этой неделе и сказал то же самое, так что я знал, чего ожидать.
«Это просто бой. С того момента, как ты киваешь головой, это просто постоянная битва все время. Ричи (Чемпион) связался с ним ранее на этой неделе и сказал то же самое, так что я знал, чего ожидать.
«Я готов к веселью. Мне нравится залезать на электронные ручки и как бы показываться и показываться на них, но в раздевалке становится более оживленно, когда возвращаются «Хопперы».
Вот что произойдет в пятницу. Всадники без седла делают небольшой перерыв после того, как садятся на самых крутых лошадей. У них будут лошади в девятом раунде, что позволит им быть немного более яркими с их шпорами. Это делает его немного веселее.
У него тоже нет времени беспокоиться о своей ничьей.Между выполнением спонсорских обязанностей и другими делами, которыми он занимается в течение дня, он также находит время, чтобы сосредоточиться на своем теле.
«Мой мануальный терапевт из дома Advantage Chiropractic & Acupuncture прилетел посмотреть несколько последних сеансов, — сказал Ларсен. «Он спустился и поработал надо мной, прежде чем я пошел на родео, и это очень помогло.
«Он спустился и поработал надо мной, прежде чем я пошел на родео, и это очень помогло.
«Постучи по дереву, у меня совсем не болит. Я первый год не болею. Я не знаю, был ли я просто готов больше, чем когда-либо прежде, но у меня никогда не было такого чувства жесткой болезненности.У нас есть шишки и синяки на этом пути, но это только часть».
Все это помогает ему показывать лучшие результаты каждую ночь, и это главная причина, по которой он участвует во второй половине чемпионата этого года.
Предоставлено twisTEDrodeo.com
NFR Прямая трансляция 2022 года Смотреть национальный финал Родео онлайн – Thalaipuseithhi
Еще одной отличительной чертой этого мягкого прожектора являются его 2 необычайно интенсивных источника света, которые могут излучать 1500 люмен.Солнце имеет 270 диапазонов и 5-летний срок службы батареи, а также светодиоды, встроенные внутрь. Мягкий фотоэлектрический поток Richarm активируется мгновенно на закате, и он фактически отключится сразу же на рассвете, если вы активируете 42 светодиодных индикатора.
Эти прожекторы работают на солнечной энергии и могут освещать дорожки, подъездные пути, пешеходные дорожки, а также внешние зоны под любым углом. К счастью, прожектор SolarBright представляет собой фотогальваническую лампу с долговечной батареей, которая работает весь вечер.Вы можете сделать установку самостоятельно, и всего за пару минут у вас уже может быть несколько ярких огней на вашем дворе.
Если вы ищете эффективное и яркое освещение снаружи, но не хотите много искать, попробуйте CLY 60 LED Photo voltaic Floodlight. Фотогальванический прожектор GLORIOUS-LITE обеспечит именно это, если вам нужен необычайно яркий свет.
Светильники на солнечных батареях являются одними из самых удобных инвестиций, которые вы могли бы сделать для своего малого бизнеса или дома.Что касается недостатков, то солнце обычно не длится 8 часов, потому что это зависит от климатической ситуации и, кроме того, от состояния фотоэлектрической панели.
Как правило, нам нужны прожекторы для укрытия или освещения места, и хотя мы не можем рисковать, что они исчезнут на нас для любого вида деятельности. Как покупатели, нам нужны лампы, которые можно использовать в течение очень долгого времени. Затем подключите прожекторы к Wi-Fi, используя прилагаемый кабель Ethernet.
Как покупатели, нам нужны лампы, которые можно использовать в течение очень долгого времени. Затем подключите прожекторы к Wi-Fi, используя прилагаемый кабель Ethernet.
Эти фотогальванические прожекторы имеют 120 светодиодов.Хрупкие датчики движения могут быть очень полезным дополнением для целей безопасности, и вы не захотите беспокоиться весь вечер, осознавая, что батарея будет стоить, когда наступит день, как только снова.
К недостаткам относится то, что солнечный свет обычно не длится 12 часов, и, кроме того, это связано с тем, что время освещения зависит от местных погодных условий, и солнечные лучи лучше всего освещают свое место. Как было указано ранее в этом обзоре, прожектор SolarBright можно оставить включенным во всех трех режимах: «Полностью вкл.», «От затемнения до сияния» или «Упражнение с блеском».
BassCliff с ANB в Gilley’s Las Vegas (NFR), 14-15 декабря 2018 г. (видео добавлено!)
Привет! Хорошо, хотите верьте, хотите нет, но в субботу вечером мы снова хорошо провели время. Всю ночь стояла очередь, чтобы попасть внутрь, но персонал там начал узнавать басиста, так что мне не пришлось платить входную плату, чтобы войти на этот раз.
Всю ночь стояла очередь, чтобы попасть внутрь, но персонал там начал узнавать басиста, так что мне не пришлось платить входную плату, чтобы войти на этот раз.
Но я забегаю вперед. После того, как мы вернулись из Sands Exposition Center, где мы делали подкаст для «Иисус зовет», я понял, что очень голоден.Было 4 часа дня, и я не ел весь день. Итак, я направился в столовую для сотрудников. У меня есть карта питания, которая позволяет мне питаться бесплатно, как и всем остальным сотрудникам.
{}Поскольку я обычно ем только один раз в день, когда нахожусь в Вегасе, я снова накормил себя белком и овощами. Рыба, сосиски, стейк Солсбери, черная фасоль, большой салат, немного брокколи на пару и одна фаршированная ракушка для макарон. Мне нужно было всего несколько углеводов.
{}Позже я подготовился к шоу и зашел в магазин в казино, чтобы забрать свою ROCKSTAR.Это не только повышение энергии, но и повышение эго.
{} Я рано добрался до хонки-тонка. Родео только что закончилось на большом экране. Там было много людей, просто чтобы посмотреть на родео.
Родео только что закончилось на большом экране. Там было много людей, просто чтобы посмотреть на родео.
Я встретил @G19Tony, и он купил мне пива, прежде чем мы начали. Спасибо, мой друг! Затем мы поднялись на сцену, чтобы позволить DJ Shaddix объявить нас.
{}Тогда мы начинаем шоу! Давай Аманда!
{}Когда Аманда поет свои дерзкие девчачьи песни, перед ней всегда танцуют несколько дерзких девчонок.
{}Рэнди скользит и скользит по стали. Арни корчит гитарное лицо. Аманда заставляет нас хорошо выглядеть и звучать лучше.
{}У Джейсона было много дел сегодня вечером. Поскольку Джонни не было сегодня вечером, он также играл партии Джонни (которые мы записали ранее) на том причудливом трековом проигрывателе Roland, который он использует.
{}Вот несколько фотографий половины группы в движении. Я уверен, что объяснения не нужны.
{} {} {} {} Было очень весело. Всю ночь мы играли только три или четыре линейных танца. Эти люди любили свои два шага и свинговые танцы. На этом снимке ди-джей играл линейный танец во время нашего перерыва. Вы можете увидеть, как народ танцует в задней части танцпола, а другие танцуют свинг в передней части зала.
Всю ночь мы играли только три или четыре линейных танца. Эти люди любили свои два шага и свинговые танцы. На этом снимке ди-джей играл линейный танец во время нашего перерыва. Вы можете увидеть, как народ танцует в задней части танцпола, а другие танцуют свинг в передней части зала.
Три волосатых мальчика на перемене.
{}Эй! Посмотрите, кто ходил на месте. Наш друг Тамми. Ты помнишь ее по концерту Swallows Inn, верно?
{}Вот еще одна моя фотография с Тамми и Рэнди, просто так.
{}Затем мы вернулись на сцену, и танцоры снова оказались на полу. Черт, но это была веселая ночь.
{}Этот концерт довольно уютный. Мы начинаем в 10 вечера и играем часовой сет, делаем получасовой перерыв, играем 45-минутный сет, делаем получасовой перерыв и играем еще 45-минутный сет, чтобы закончить ночь в 13:30. Вот банда начинает собираться.
{} Я отключил свою бас-гитару, положил ее в футляр и ушел оттуда примерно через 5 минут. Мне нравится быть басистом. Я даже не привожу на этот концерт усилитель, только IEM и бас. Я подключаюсь к домашнему DI-блоку и готов к работе. Звукорежиссеры любят меня, потому что им никогда не приходится возиться с басом. Я делаю проверку звука одной ноты, и я готов к работе.
Мне нравится быть басистом. Я даже не привожу на этот концерт усилитель, только IEM и бас. Я подключаюсь к домашнему DI-блоку и готов к работе. Звукорежиссеры любят меня, потому что им никогда не приходится возиться с басом. Я делаю проверку звука одной ноты, и я готов к работе.
В любом случае, все, что осталось, это ехать домой, что всегда занимает больше времени, чем дорога сюда. По крайней мере, я смогу посмотреть на пустынный пейзаж по дороге домой. Спасибо всем, кто делится с нами своими выступлениями здесь.Спасибо, что посмотрели на меня. У меня будет пара выходных, а потом мы будем играть в Stillwater Spirits and Sounds в Дана-Пойнт. До скорой встречи.
Спасибо за снисходительность,
BassCliff
Кураторы Дубии беспокоятся о пленках Никсона
Кампания Джорджа Буша-младшего выразила озабоченность по поводу публикации 420 часов секретных записей Белого дома бывшего президента Никсона. На пленках есть голос отца кандидата в президенты, бывшего президента Джорджа Буша.
Джордж Буш разговаривает или о нем говорят в более чем 100 из 4140 разговоров, которые должны быть обнародованы Национальным архивом. На пленках записаны разговоры конца 1971 года, когда бывший президент был послом США в ООН.
«Мы действительно звонили, чтобы выразить беспокойство», представитель кампании Буша Ари Флейшер заявил в среду вечером. «У нас есть постоянная озабоченность по поводу того, собирается ли эта администрация создавать внешние события, чтобы повлиять на исход этих выборов.»
«Мы выразили озабоченность по поводу сроков. Нам сказали, что это заранее определенные даты (для выпуска магнитофонных лент Никсона), и мы это принимаем», — сказал Флейшер.
В четверг архивы планируют обнародовать более 4000 разговоров, записанных Никсоном в период с августа по декабрь 1971 года. Ключевые республиканские фигуры, которые могут быть на записях, могут повлиять на кандидатуру его сына на пост президента всего за две недели до выборов.
Джон Х. Тейлор, исполнительный директор Фонда библиотеки и места рождения Ричарда Никсона в Йорба Линда, Калифорния, знаком с выпускаемыми пленками Никсона.
«Я в значительной степени знаком с содержанием выпуска. Я могу заверить вас, что нет абсолютно ничего, что нанесло бы ущерб семье Бушей», Тейлор.
Предвыборный штаб Буша подверг резкой критике членов кабинета президента Клинтона за то, что они поддержали предвыборную кампанию демократа Эла Гора.
«Мы обеспокоены тем, что эта администрация делает что-либо, что может дискредитировать республиканцев», — сказал Флейшер.
Флейшер сказал, что кампания выразила свою обеспокоенность Джиму Чиккони, который работал в Белом доме Буша и помог создать президентскую библиотеку Буша. Затем Чиккони позвонил директору Национального архива Джону Карлину, чтобы узнать о выпуске ленты Никсона, сказала Сьюзан Купер, пресс-секретарь архива.
Она сказала, что не знает, просил ли Чиккони архивариуса отложить публикацию записей до окончания выборов.
Никсон установил свою систему записи в Белом доме в начале 1971 года. Существование системы записи было раскрыто во время дачи показаний в Уотергейтском комитете Сената в 1973 году. Правительство конфисковало записи, когда Никсон ушел в отставку. После долгих лет судебных тяжб Национальный архив начал выпускать партии записей, о которых раньше никто не слышал.
CBS Worldwide Inc. Все права защищены. Этот материал нельзя публиковать, транслировать, переписывать или распространять.Ассошиэйтед Пресс способствовало этому отчету
Актуальные новости
Загрузите наше бесплатное приложение
Для последних новостей и анализа Загрузите бесплатное приложение CBS News
.